A Neurophysiology and Clinical Reasoning Approach to Dosage and Manual Therapy
Martin Krause
Bachelor of Applied Science (Physiotherapy), University of Sydney
Masters of Manipulative Physiotherapy, University of Sydney
Graduate Certificate Health Science Education, University of Sydney
Graduate Diploma Health Science (Exercise and Sport), University of Sydney
Certificate IV Workplace Assessment and Training
Private Practitioner, North Sydney, Australia.
Table of Contents
Neurogenic and non-neurogenic inflammation
Sympathetic modulation of inflammation and it's role in repair
The consequences of reduced receptor threshold to manual therapy
Secondary mechanical hyperalgesia due to oedema around the dorsal root ganglion
Spinal cord expansion of sensitised receptive fields
Manual therapy during peripheral and central expansion of the hypersensitive receptive field
Introduction
Pain is a dynamic entity, which affects a large number of people around the world. Pain knows no cultural boundaries and people suffering from pain have been treated in various manners by various practitioners with various degrees of success for centuries. Modern medical and paramedical practice has attempted to come to terms with patient's ailments by diagnosing various pathologies with labels such as 'fibromyalgia', 'fibrocytis', 'repetitive strain injury', 'cervico-brachialgia', 'reflex sympathetic dystrophy', etc. However these labels may detract from effective clinical reasoning because
- the diagnostic criteria become so broad that it becomes difficult to exclude people from that diagnosis (Cohen & Quintner 1993; Quintner & Cohen 1994).
- the fundamental clinical reasoning principle for the production of symptoms is violated. i.e. a leap is made from signs and symptoms (phenomenology) to the assumption of disease (nosology) without consideration of pathophysiology (Cohen 1995).
To improve clinical reasoning skills it may be important for the clinical practitioner to understand the pathophysiology that underlies the phenomenology (Jones 1995). It is the objective of this paper to elucidate the mechanisms of pain and inflammation in a theoretical and clinical context. Improved understanding of pathology may provide the practitioner the context specific feedback to discover the most effective approach to treatment. Hence, the main argument which will be pursued here is that clinical signs and symptoms correlate with the underlying process of inflammation and repair. Thus, the dossage and type of manual therapy employed in the treatment of pain should reflect the normalisation of clinical signs and symptoms.
Inflammation
The reaction of living tissue to injury is called inflammation. Inflammation involves a cascade of events in cells, involving products of cells, enzymes, soluble factors, and blood vessels and their contents (Enwemeka & Spielholz 1992; Schmidt et al 1994; Tillman & Cummings 1992). Trauma to the intervertebral disc (IVD) may result in a focus of inflammation inside the disc (Weinstein et al 1988). Inflammatory substances may diffuse out of the IVD and contact the posterior longitudinal ligament, Batson's venous plexus, Hoffman ligaments and the dura mater (Adams et al 1986; Kambin et al 1980; Liu et al 1991; MacMillan et al 1991; Troisier & Cypel 1986). In contrast to the IVD, these later structures have a good blood supply and are densely innervated by sensory and sympathetic nerve fibres. Consequently, the nerve receptors and blood vessels may become inflamed directly from trauma and/or indirectly from inflammatory substances of IVD trauma (Korkula et al 1985; McLain 1993; Weinstein et al 1988; Wiltse et al 1993). Hence, inflammation and the resolution of inflammation may be synonymous with healing.
Inflammation and oedema
Inflammation may cause the formation of oedema and spinal pain (Barker et al 1991; Gallin et at 1992; Groenblad et al 1991;1994b; Kawakami et al 1994a; 1994b; McKenzie & Saunder 1990; Rothwell & Hopkins 1995; Rydevik et al 1989; Williams & Hellewell 1992). As oedema increases, compression of intraneural blood vessels and nerve fibres may preferentially decrease the conduction velocity of myelinated afferents and efferents, whilst probably leaving the conduction velocity along unmyelinated nerve fibres largely unaffected (Cornefjord et al 1992; Kobayashi et al 1993; Matsui et al 1992; Olmarker et al 1989; Rydevik et al 1984; Takahashi et al 1993). Decreased conduction along large diameter myelinated (type II) nerve fibres has been postulated to decrease the modulation (by wide dynamic range neurones [WDR]) of nociception in the spinal cord and thus increase pain (figure 1) (Dickenson & Sullivan 1987; Ochoa & Yarnitsky 1993; Price et al 1994).
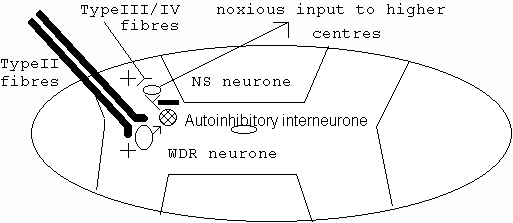
Figure 1: Spinal cord modulation of pain. Wide dynamic range (WDR) neurone inhibition of nociceptive specific neurone (NS) in the dorsal horn of the spinal cord (adapted from Wall & Melzack 1989, pp11-13; Bogduk 1993, p59)
By implication, non-noxious mechanical stimuli from joint mobilisations may be ineffective where a decreased conduction along large diameter myelinated (type II) nerve fibres exists. Hence a more effective means of treatment may be one which normalises blood flow to neural tissue (eg. traction or a sustained rotatory technique) .
Neurogenic and non-neurogenic inflammation.
Inflammation involves a complex cascade of reactions at the peripheral receptor terminals. These reactions may be divided into two entities: neurogenic and non-neurogenic inflammation. Neurogenic inflammation includes the release of neuropeptides (e.g. substance P, Calcitonin Gene Related Peptide [CGRP], nordrenaline, etc) from C-fibre terminals whereas non-neurogenic inflammation involves the release of inflammatory substances (e.g. histamines, prostaglandins, cytokines, leukotrienes, bradykinin, etc) from the blood vessels and connective tissue. Both processes lead to a decrease in the activation threshold of the mechanoceptors.
Neurogenic inflammation is the process whereby neuropeptides released from C-fibres of primary afferents and sympathetic efferents cause vasculature membrane changes which results in the leakage of proteins from the vasculature into surrounding tissue (Ahmed et al 1991; Blottner & Baumgartner 1994; Donnerer et al 1991; Heller et al 1994; Kobayashi et al 1993; LaMotte et al 1991; Levine et al 1986; Markowitz et al 1989; Suval et al 1987; Zochodne 1993). In the absence of inflammation, sympathetic nerve fibre efferent terminals in experiments with rats (Hu & Zhu 1989) suppress C-fibre afferent receptor evoked discharges. However, during inflammation C-fibre receptors increase their neurogenic inflammatory response and hence increase their discharge frequency with stimulation of the sympathetic nerve terminals (Levine et al 1986). Sensitisation of peripheral nerve terminals by neurogenic and non-neurogenic inflammation may reduce mechanoceptors thresholds and contribute to pain (figure 2 and 3) (Groenblad et al 1991; Jaenig 1985; Lotz et al 1988; Handwerker et al 1991; Levine et al 1986a; Rees et al 1994; Raja et al 1988; Rothwell & Hopkins 1995; Schmidt et al 1994; Taiwo & Levine 1989).
Massive infiltration of macrophages into the dorsal root ganglion (DRG) was demonstrated in both the ipsilateral and contralateral lumbar but not thoracic DRG using the rat model of unilateral antigen-induced arthritis (AIA) in the knee (Segond von Banchet et al 2009). This activation wasn't explainable as a result of nerve cell lesion. During AIA lumbar but not thoracic DRGs exhibited bilateral de novo expression of vascular adhesion molecule (VCAM-1) which is known to be involved with macrophage infiltration. Chemical inhibtion of tumor necrosis factor alpha after induction of AIA significantly inhibited macrophage infiltration as well reduced the expression of VCAM-1. This also reduced mechanical hyperalgesia at the inflammed joint without attenuating the joint inflammation, additionally the mechanical hyperalgesia in the contralateral knee was also reduced. Thus, bilateral segment specific infiltration of macrophages into the DRGs is part of an unilateral inflammatory process in peripheral tissue and may be involved in the generation of peripheral mechanical hyperalgesia in the non-inflammed side (Segond von Banchet et al 2009).
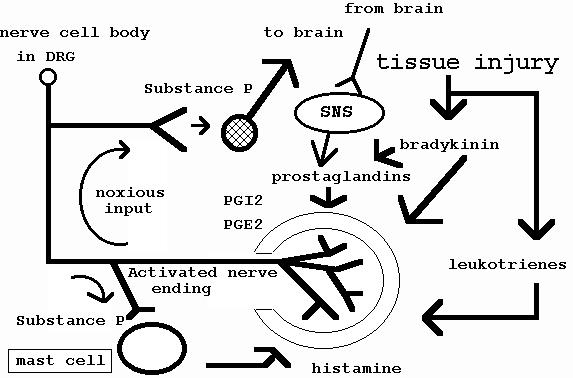
Figure 2 : Schematic representation of the interaction between neurogenic and non-neurogenic inflammation on free nerve endings through the stimulation of mast cells. SNS = sympathetic nervous system. (adapted from Groenblad et al 1991, p617; Jaenig 1985; Lotz et al 1988; Handwerker et al 1991; Levine et al 1986a; Taiwo & Levine 1989).
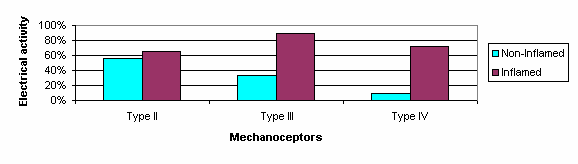
Figure 3 :Percentage of mechanoceptors (group II, III, IV) responding to movement in the non-inflamed and inflamed states (Raja et al 1988; p576)
Sympathetic modulation of inflammation and its role in repair.
The sympathetic postganglionic nerves release inflammatory mediators that increase plasma extravasation, including prostaglandins (Coderre et al 1989; Gonzales et al 1991; Gonzales et al 1989; Green et al 1991). Additionally, under conditions of sympathetic postganglionic neuronal modulation bradykinin has been found to increase plasma extravasation (Green et al 1992). The sympathetic postganglionic neurones also release mediators which decrease plasma extravasation, including neuropeptide Y and noradrenaline (Green et al 1991;1992). Apart from modulating plasma extravasation these substances may also interact with endothelial relaxing factor (Greenberg et al 1991) and platelet-activating factor (Heller et al 1994) in the blood vessels. Agents that enhance synovial plasma extravasation have been demonstrated to decrease tissue injury during inflammation (Coderre et al 1991). The mechanisms by which sympathetic nerve terminals decrease tissue injury are unclear. However, an increased plasma extravasation would be expected to reduce the concentration of inflammatory substances through the facilitation of lymphatic and venous drainage, thereby improving repair (figure 4)(Heller et al 1994).
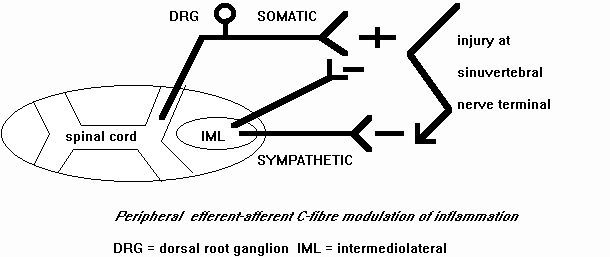
Figure 4 : Sympathetic modulation of peripheral nerve activity
The consequences of reduced receptor threshold to manual therapy.
At this stage it would appear that there are 2 mechanisms of modulation of electrical activity in type III and IV nerve fibres. These 2 mechanisms include
- non-noxious stimuli via the type II nerve fibres and WDR neurones and
- the peripheral sympathetic nervous system.
Significantly, WDR neurones probably respond specifically to the wide range of frequencies of input from the variations of force, frequency, direction and velocity which gentle oscillatory mobilisation techniques present. Consequently the type and dosage of techniques used need to reflect the normalisation of signs and symptoms. Further, the SNS is likely to respond to techniques which increase blood flow and lymphatic drainage whilst simultaneously being demonstrated to be non-threatening and effective to both the patient and the therapist. Thus, after each treatment technique the ‘expected' normalisation of signs and symptoms need to be demonstrated. Similar to the paradox of Schroedingers Cat in quantum physics, it is not until we observe an effect do we know the effect. Yet our mere observation can affect clinical outcomes.
Secondary mechanical hyperalgesia due to the effect of oedema on the nerve roots/dorsal root ganglia/spinal nerve complex.
Endoneurial oedema of the dorsal root ganglia (DRG) may be the result of compression from inflammatory exudate (Chatani et al 1995). Endoneurial oedema of the DRG may generate ectopic electrical impulses (Bandalamente et al 1987). Ectopic impulses originating in the DRG are thought to propagate into the spinal cord and into peripheral receptor sites (Bandalamente et al 1987; Wall & Devor 1983). Increased neuronal firing may increase neurogenic inflammation at the receptor sites (Chatani et al 1995; Markowitz et al 1989; Xavier et al 1990). Arborization of peripheral nerve terminals may result in neurogenic inflammation in an expanded receptive field (LaMotte et al 1991). This suggests that extraneural inflammation may induce intraneural oedema that in turn may increase the intensity of extraneural neurogenic inflammation. In the case of the sinuvertebral nerve this may mean that receptors at multiple (eight) segments may become sensitised and exhibit decreased mechanical activation thresholds (Groen et al 1988; Koltzenburg et al 1994; Raja et al 1988). This area of neurogenic inflammatory expansion of the sensitised receptive field is termed secondary mechanical hyperalgesia (Raja et al 1988).
During surgery, retraction of the inflamed dura mater has been demonstrated to increase nerve impulse generation (Greenbarg et al 1988; Kuslich et al 1991). Therefore, the clinical implications of decreased activation threshold are a decreased dosage of treatment. Feedback of the effect of treatment on signs and symptoms may be a means of determining the effective dosage of treatment.
Interestingly, a projection of nerve fibres from the DRG to the pia mater of the ventral nerve root has been demonstrated in the cat (Jaenig & Koltzenburg 1991). Since electrical stimulation of the sympathetic nerve fibres has been demonstrated to sensitise mechanoceptors in rats (Barasi & Lynn 1986; Roberts & Elardo 1985; Snajue & Jun 1989), then ectopic impulse propagation due to mechanical irritation of the DRG may result in neurogenic inflammation of the ventral nerve root pia mater (Groenblad et al 1991; Harvey et al 1994; Wall & Devor 1983). However, the effect of neurogenic inflammation of the ventral nerve root on sympathetic b -fibre and a -motor fibre activity is theoretically and clinically uncertain.

Spinal cord expansion of sensitised receptive fields.
Spinal cord sensitisation (hypersensitivity) of adjacent wide dynamic range (WDR) and nociceptive specific (NS) neurones may occur due to the direct synaptic connections between adjacent WDR neurones and between adjacent NS neurones (figure 5)(Price et al 1994). Spinal cord expansion of the sensitised (hypersensitive) receptive field has been demonstrated and is thought to be a compensatory mechanism for the dissipation of the afferent barrage of electrical stimulation from sensitised (hypersensitive) peripheral receptors (Coghill et al 1991; LaMotte et al 1991; Magal 1990; Mao et al 1992; Price et al 1994; Torebjoerk et al 1992; Vaccarino et al 1987) and from ectopic impulse generation from the DRG (Wall & Devor 1983). Evidence for central expansion of the neuronal receptive field comes from the findings that if the nerve of the new hypersensitive receptive field of the adjacent dorsal horn neurones is ligated no expansion of the sensitivity of the peripheral receptive field occurs (Attal et al 1994). This suggests that in the presence of inflammatory exudate around the nerve root/DRG/spinal nerve complex and/or the sinuvertebral nerve then an adjacent peripheral nerve (e.g. sinuvertebral nerve, median nerve, etc,) may be recruited for the projection of neurogenic inflammation into adjacent areas of the spinal canal and/or limb.

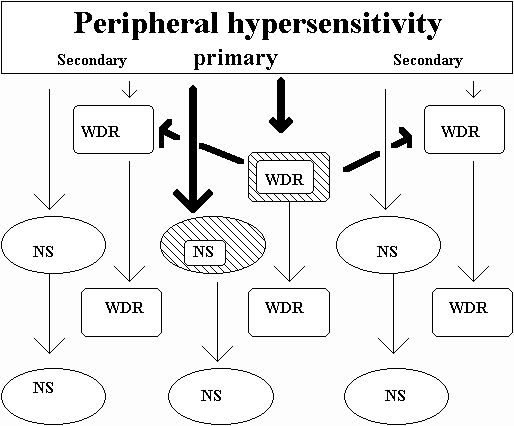
Figure 5: Central expansion of the hypersensitive receptive field.
The significance of treating the pain with manual therapy during peripheral and central expansion of the hypersensitive receptive fields.
Both people in pain and therapists may have varying difficulty in localising the source of symptoms. Significant expansion of the hypersensitive receptive field may be postulated to result in increased acuity to detect the locus of dysfunction through a "3 dimensional map" of areas of normal activity and those of primary and secondary hyperalgesia. Perhaps the analgesic effect of gentle stroking around a painful area may be an example of this mechanism? Hence, in the case of referred neurogenic pain non-noxious manual therapy directed at the site of pathology may enhance the modulation of electrical activity in type III and IV fibres through mechanical input to aid in the detection of the locus of dysfunction.
Alternatively, expansion of the hypersensitive receptive field may be postulated to result in decreased ability to detect the locus of dysfunction. A clinical example of such a scenario may be when referred limb pain is more intense then the spinal pain at the site of pathology (eg spinal nerve/dorsal root ganglion/nerve root irritation). In this example it is highly probably that distal limb pain can be more severe than proximal pain due to the high density of mechanoceptors in the hands and the feet and their resultant disproportionate representation in the somatosensory cortex. (Interested readers should refer to somatosensory homunculus for further details). Therefore, in the case of projected neurogenic limb pain it may be hypothesised that unloading the site of secondary neurogenic inflammation (with for example tape) may increase the awareness of the primary site of pathology. Regardless of the effect of central expansion of the hypersensitive receptive field, it is highly likely that determination of the precise source of symptoms is necessary for effective healing to take place.
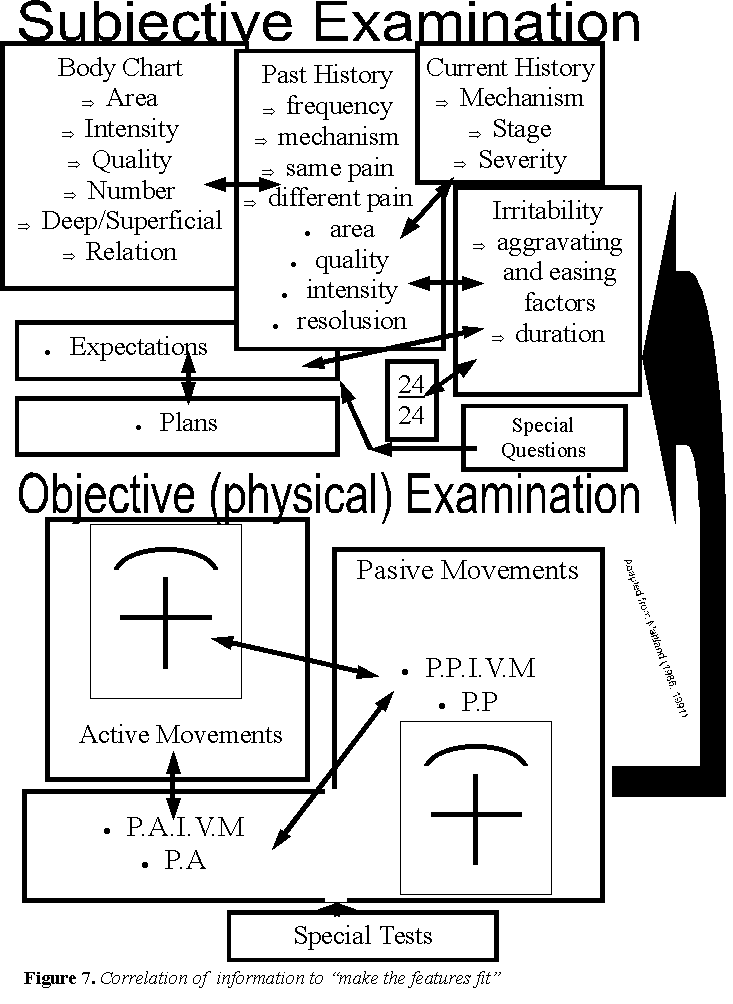
Clinically, the subjective examination includes a body chart which describes the area of pain. Hence, pain in a dermatological distribution which may be accompanied by pins & needles and numbness should result in the practitioner using a different line of clinical reasoning then that for referred somatic pain. Therefore, inductive questioning should reflect the responses from deductive questioning (figures 6 & 7).

Figure 6 : Reasoning involved differentiating Somatic from Radicular pain

Decreased movement due to the effect of expansion of spinal cord neuronal and peripheral receptor hypersensitivity associated with spinal and limb pain.
Several experiments have been designed to differentiate the contribution of peripheral receptors and spinal cord neuronal plasticity in pain production. Expansion of the hypersensitive receptive field (Price et al 1992) together with the decreased activation threshold of low and high threshold mechanoceptors (Ferrell et al 1988; Koltzenburg et al 1994; Raja et al 1988; Schmidt et al 1994) and the five-fold increase in resting discharge of an articular nerve from an inflamed joint (Schmidt 1990) would mean that spinal cord neurones receive an unusually large barrage of nociceptive inflammation from inflamed deep tissue. These are the conditions thought to be required for intracellular changes in WDR neurones that would make them hyperresponsive (Dickenson & Sullivan 1990; 1987; Thompson & Woolf 1991; Urban & Randic 1984). Investigations demonstrate that if a source of ongoing nociceptor input continuously "refreshes" the hyperexcitable state of the dorsal horn neurones (Gracely et al 1992) then this may lead to the loss of function of the auto-inhibitory inter-neurones between the WDR and NS neurones (Alkon &Rasmussen 1988; Collingride & Singer 1990; Laird & Bennett 1993; Nishizuka 1989). Clinically, hyperexcitability of WDR neurones may result in motor abnormalities such as the marked and prolonged increase in the flexion withdrawal reflex, as exhibited in rats (Ferrell et al 1988; Woolf 1983; Woolf et al 1994). The rat will try to avoid contact of the inflamed paw with the ground (Woolf 1984; Woolf & McMahon 1985; Woolf & Wall 1986). A similar situation may arise in humans whereby joint movements are prevented by increased muscle activity so as to minimise the mechanical input onto inflamed mechanoceptors. Lack of movement may be postulated to effect blood flow with a resultant exacerbation of oedema and/or ischaemia. Furthermore, lack of movement may be postulated to result in decreased non-noxious mechanical input that may be required for the modulation of electrical activity in both WDR and NS neurones. Therefore, lack of movement is likely to result in pain.
Further, more potent neurotoxic effects on autoinhibitory interneurones may occur due to ectopic input onto WDR neurones from a nerve subject to constriction (Dubner 1991; Sugimoto et al 1990). Animal models using ligatures around the sciatic nerves demonstrate large increases in spontaneous discharges in large and small diameter myelinated afferents, whilst leaving small diameter unmyelinated afferents unaffected (Bennett & Xie 1988). Thus, inflammation within the nerve itself and at peripheral nerve terminals in the limb and/or around the nerve itself may contribute to decreased modulation of pain at the spinal cord level. Conceivably, therapeutic intervention which decreases the mechanical irritation, normalises blood flow and also activates the sympathetic nervous system's (SNS) neuromodulation of inflammation, muscle activity and blood flow may also activate WDR modulation of NS neurones and hence decrease pain.
The effect of manual therapy on muscle activity around the intervertebral foramen (IVF).
Manual therapy may be directed at reducing muscle spasm. It is conceivable that sustained techniques may have a direct effect on the stretch reflex of muscles. Alternatively, a sustained technique (e.g. traction) may decrease electrical activity in type III and IV fibres through normalisation of blood flow and decreased mechanical pressure generated by inflammation on soft tissue structures in and around the IVF. Indeed, only 30mmHg pressure is required to obliterate endoneurial venous return (Lundborg 1988). Finally, non-noxious input from gentle oscillatory pressures may generate sufficient WDR modulation of NS neurones to allow decreased muscle spasm and increased functional muscle activity to take place. Any decrease in pain should be followed by functional exercises to maintain soft tissue integrity. Further, examination of the neurological signs and symptoms, and the painful active movements should be examined to demonstrate the effects of treatment.
The dosage of manual therapy and the significance of the 'stage' of inflammation and 'irritability' on ‘24 hour behavior', ‘current' and ‘past history'.
The stage of the inflammation and the ‘irritability' (Maitland 1986; 1991) may influence the treatment approaches under consideration. For example, in an ‘irritable' dysfunction and/or during the acute stage of injury the non-neurogenic inflammatory process may be more beneficially affected by sustained mobilisation techniques or constant traction which are thought to relieve the pressure around the nerve tissue. If this hypothesis were clinically true, then neurological changes such as reduced muscle power, reduced reflexes and reduced sensation should improve if the correct technique (position and amount of force) are used. During the later stages of inflammation and/or in a ‘non-irritable' dysfunction the clinical signs and symptoms may be more readily influenced by treatments (e.g. joint mobilisations, mobilisations of the SNS, taping) specifically directed at the neurogenic inflammation. Treatment directed at the neurogenic inflammation must ascertain the status of the receptor thresholds with the aim of treatment being a decrease of afferent electrical activity through an increase in the modulation of the receptor thresholds of the type III and IV mechanoceptors. The principles of these hypotheses involve the fact that if inflammation results in a decrease in the receptor thresholds and a decrease in the modulation of electrical activity in type III and IV fibres, then the reverse should be true as inflammation resolves. That is, receptor thresholds and modulation of afferent electrical activity should increase. The expected result is decreased pain and consequent increases in ranges of movement, normalisation of voluntary muscle power, sensory discrimination and reflexes. Improvement in the clinical status of the patient should simultaneously correlate with improvements in the ‘irritability' and ‘24 hour behaviour' of the dysfunction. In this manner the practitioner must treat the pain and not just any pain of which the patient complains (figure 7).
If oscillatory mobilisation techniques are employed at a ‘stage' too early during the inflammatory process (i.e. in a worsening condition [e.g. distal pain > proximal pain] and/or in an ‘irritable' dysfunction and/or progressing ischaemic compromise of a nerve root/DRG/spinal nerve), then this may exacerbate muscle spasm directly due to a stretch reflex or indirectly due to activation of inflamed receptors and neural compromise. Summation of afferent electrical activity from inflamed receptors may result in pain and further 'splinting' of the joint. Under no circumstance should an examination process or technique be used that increases neural compromise. Hence specificity of dosage is crucial not only for optimal results but also for safety.
Importantly, the clinician will need to ascertain the expectations of treatment outcome in relation to the ‘stage' and ‘irritability' which may be questioned in the subjective examination through not only the ‘24 hour behaviour' but also through ascertaining the ‘current' and ‘past history' of the signs and symptoms. In this manner improvements or deterioration in the patient's condition may be monitored and attributed to either the consequences of the type and dosage of treatment or to the consequences of the natural progression of the pathology.
Descending inhibition in the modulation of pain.
It is quite likely that the sympathetic nervous system (SNS) is involved in very specific but very different ways depending on the stage of inflammation and the ‘irritability' of the dysfunction. The peripheral modulation of inflammation by the SNS (discussed previously) may be quite distinct and more readily influenced by enhanced blood flow, reduced oedema and reduced muscle spasm due to movement of inflammatory exudate. However, in presence of reduced autoinhibitory interneurone modulation of pain (in the spinal cord by WDR neurones on NS neurones) then at least two potential sources of descending neuromodulation may be recruited. One source of descending inhibition involves pontine noradrenergic projections onto ventral and dorsal horn neurones involved with the modulation of pain (figure 8) (see Proudfit 1992 for review; Jaenig 1985; Morgan et al 1989; Nakagawa et al 1990; Post et al 1986; Ren et al 1990). Descending noradrenergic inhibition is considered to be an opioid-independent form of analgesia (Proudfit 1992) however, evidence exists which supports the view that m -opioid and a -2 noradrenergic receptors are functionally linked to pain modulation (Kalso et al 1993). Apart from animal investigations for descending sympathetic pain modulation, further evidence comes from clinical investigations where the sensory and affective aspects of ischaemic pain (Maixner et al 1990) and muscle spasm (Shindo et al 1994) correlate with SNS induced cardiovascular responses.
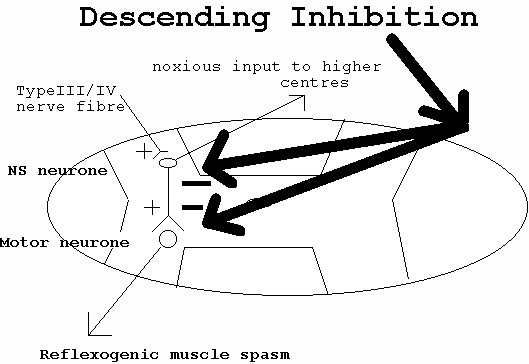
Figure 8 : Descending modulation of pain (adapted from Wall 1995)
Manual therapy and descending modulation of pain.
Investigations of grade III PA oscillatory mobilisation (Maitland 1986) of the cervical spine (C5/6) demonstrated an initial sympathoexcitatory effect which started very rapidly (within 15 seconds) after the commencement of treatment (Petersen et al 1993). Wright & Vicenzino (1995) attributed the modulation of the sympathoexcitatory effect on the dorsal periaqueductal grey (dPAG) via nucleus paragigantocellularis, using noradrenaline as the transmitter substance. A review (by the investigators) of the acupuncture literature suggested that this initial sympathoexcitatory effect might be followed by sympathoinhibition some 20-45 minutes later (Wright & Vicenzino 1995). This sympathoinhibition was postulated to be as a result of modulation by the ventral PAG (vPAG) via the nucleus raphe magnus (NRM) to the dorsal horn and intermediolateral horn (IML) of the spinal cord to effect analgesia and cardiovascular changes. The connection from the nucleus raphe pallidus and obscurus to the IML and anterior horn is thought to influence motor activity and autonomic function (Lovick 1991, cited by Wright & Vicenzino 1995). These preliminary results are interesting, as there are very few adequate studies that have investigated the neurophysiology of the analgesic effects of manual physiotherapy.
Sensorimotor "center-surround inhibition" of pain.
Apart from the SNS, another source of descending modulation of pain is considered to arise from the sensorimotor cortical regions involved with input on spinal cord and brainstem regions (Hsieh, Stahle-Backdahl, Hagermark, Stone-Elander, Rosenquist, Ingvar 1995; Wall 1995). Wall (1995) describes this system as setting a "sensory posture" whereby motor input exerts 'powerful' control over afferent spinal cord processing which 'focuses on action rather than perception' (p35) (also Galea & Darian-Smith 1995). Wall (1995) used positron emission tomography (PET) studies in patients with chronic pain to substantiate this claim whereas Galea & Darian-Smith (1995) used PET and anatomical dissection of primates. Further evidence for descending modulation comes from Gogas et al (1991) where supraspinal opioids were found to block the perception of pain through a 64% reduction in superficial laminae dorsal horn activity (usually associated with a predominance of nociceptive specific [NS] neurones (Lima et al 1994)) and an 85% reduction in ventral horn activity (usually associated with motor activity) (Schomburg & Steffens 1991). Investigations substantiated the finding that higher centers can act in an antinociceptive manner where a predominance of inhibition occurs at NS neurones but where excitation or inhibition occurs with equal frequency on wide dynamic range (WDR) neurones (Sandkuehler et al 1995). This appears to be a unique way of looking at pain perception where the cortical input is primary to a secondary ascending nociceptive stimulus. This arrangement is postulated to improve discrimination of NS neuronal activity and thereby possibly decrease pain (figure 9) (Bogduk 1993; Laird & Cervero 1990).
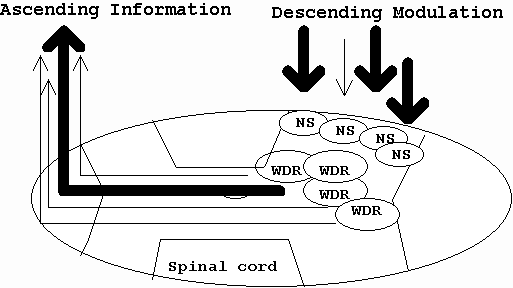
Figure 9 : Centre – Surround Inhibition (Bogduk 1993)
The focus on action with functional tape and motor control in the treatment of pain.
The clinical application of functional tape and specific motor control suggest a marked pain inhibitory effect beyond that which may be explained purely on biomechnical grounds. Since primary hyperalgesia in deep tissue may lead to secondary hyperalgesia in superficial structures then therapeutic intervention, which results in a decrease in secondary hyperalgesia, may be postulated to result in increased movement.
Evidence for descending inhibition comes from a recent study on the modulation of cutaneous nociception concluded that besides the brainstem, other higher regions may be responsible for the marked descending inhibition of nociception (Hsieh et al 1995; Morton, Siegel, Xiao, Zimmermann 1997). Specifically, the somatosensory-motor cortex has been postulated to be involved in movement dysfunction as a consequence to and as a consequence of pain. It is conceivable that the cutaneous feedback from tape and the decrease in noxious afferent input may increase the discriminatory role of descending inhibition. Further, the specificity of motor control may also increase the descending inhibition of nociception.
Evidence for the effect of pain on the specificity of motor control comes from investigations into low back pain. Altered patterns of abdominal muscle activation with arm or leg movements have been recorded in patients with chronic low back pain (O'Sullivan, Twomey, Allison, Sinclair, Miller, Knox 1997; Hodges and Richardson 1996; Hodges, Richardson, Jull 1996; Richardson and Jull 1995). Therefore, the role of specific exercise, specific functional tape, and specific feedback may be to modulate pain and improve muscular co-ordination.
Apart from tape and specific exercise, feedback may come in the form of demonstrating improvements in the signs and symptoms both to you and the patient. Demonstrating improvements in the symptoms may be possible only through a very discriminative subjective examination (figure 7). A discriminative subjective examination may use the Maitland (1986, 1991) approach as well as pain questionnaires such as McGill, Oswestry, etc. Further, the improvements in the clinical signs may occur through range of movement testing, pressure biofeedback, EMG biofeedback and the testing of neurological signs and neurological tension tests. Finally the ability of the manual therapist to discriminate the site of pathology in the spine has been demonstrated to be valid and reliable and to have high inter-examiner reliability (Jull, Zito, Trott, Potter, Shirley, Richardson 1997). This means that both the therapist and the patient are able to judge the effect of treatment technique and dosage on the current ‘stage' of the dysfunction if an organised assessment and treatment procedure is implemented.
Conclusions: The need for a thorough subjective and physical examination to be able to make clinical decisions, to apply the correct technique and dosage, and to assess the expected treatment outcome.
The clinical implications of descending modulation of pain suggest that during the application of manual physiotherapy to the cervical spine a noticeable treatment effect may occur within 15 seconds of mobilisation. Furthermore, it would appear that if mobilisation does result in a focus by the patient on 'action', then it is imperative to re-examine the active movement (incl. combined active movement) which initially reproduced the patient's pain.
Since descending modulation involves a principle of "fine tuning" of the perception of pain, then the initial examination must be very discriminative as to the exact area and range of movement where pain occurs during active movements. Also the ‘quality' and ‘intensity' of pain and the ‘relationship' between the pains must be noted since secondary mechanical hyperalgesia and spinal cord hypersensitisation may occur as a result of neurogenic inflammation.
Movement aberrations/inco-ordinations due to muscle spasm and/or due to muscle weakness, decreased reflexes and sensation should be noted and correlated with known anatomical innervations.
Meticulous recording of information may allow the practitioner to use their cognitive processing for analysis rather than for remembering. By implication, the questioning of the patient through the clinical reasoning process may be hypothesised to also aid the patients' cognitive processing and hence modulation of pain.
The information from the examination of active movements should correlate with the ‘aggravating/easing factors' and ‘24 hour behaviour' of the pain as well as with ‘stage', ‘stability' and ‘irritability' of the dysfunction. Only with a discriminative subjective examination will it be possible for the practitioner to focus their clinical reasoning skills in search of the pain (not any pain) during their physical examination. However the practitioner may never know the effect of treatment if the initial examination has left the practitioner with nothing on which the practitioner nor the patient can judge the expectations of treatment outcome (see Mechanical traction - clinical example of neurophysiological and biomechanical pain modulation). Finally, visualisation, verbalisation and goal orientation have powerful influences on motor learning. Improved motor performance reduces the likelihood of re-injury through increased awareness of the correct patterning of movements (see Motor Learning and Orienteering expertise) . This, in turn will improve the force closure required for stability around a joint (see Pelvic Girdle Pain - muscle energy techniques - Swiss Ball).
Many dysfunctions will improve with time, however the discriminatory practitioner should be able to predict treatment outcome and either minimises the period of pain and/or prevent an acute injury from becoming chronic by applying the optimal dosage of treatment.
Conceptualised and written by Martin Krause for lectures presented between 1997 and 1999. Since this time an overwhelming body of evidence supports the neurogenic modulation of pain and inflammation in cortical, subcortical, cerebral, spinal cord, dorsal root ganglion, peripheral sympathetic nervous system and afferent somatic nervous system sites. Undoubtedly, the cortical involvement highlights the importance of explanations and education of your patient during treatment. Feedback, both visual and verbal, as well as realistic goal setting appear to be of paramount importance for involving the patient in their recuperation (Martin Krause 2004).
- Motor Learning and gaining of expertise through avoidance of injury
- Contemporary approach to Manual Therapy
- Motor control of the cervical and lumbar spine
- Psychosocial and cognitive aspects of pain
- Instability of the Cervical and Lumbar Spine
- Chronic Low Back Pain
- Immune Responses
- Pain in the Brain
- Fibromyalgia
- Stability
Clinical Examples
Adams, M.A., Dolan, P.& Hutton, W.C. (1986). The stages of disc degeneration as revealed by discogram. Journal of Bone and Joint Surgery 68B , 36.
Ahmed, M., Bjurholm, A., Kreicbergs, A.& Schultzberg, M. (1993). Sensory and autonomic innervation of the facet joint in the rat lumbar spine. Spine 18 (14), 2121 to 2126.
Alkon, D.L.& Rasmussen, H.A. (1988). A spatial-temporal model of cell activation. Science 239 , 998 to 1005.
Attal, N., Filliatreau, G., Perrot, S., Jazat, F., Di Giamberardino, L.& Guilbaud, G. (1994). Behavioural pain-related disorders and contribution of the saphenous nerve in crush and chronic constriction injury of the rat sciatic nerve. Pain 59 , 301 to 312.
Badalemente, M.A., Dee, R., Ghillani, R., Chien, P-F. & Daniels, K. (1987). Mechanical stimulation of dorsal root ganglia induces increased production of substance P : A mechanism for pain following nerve root compromise? Spine 12 , 552 to 555.
Barasi, S.& Lynn, B. (1986). Effects of sympathetic stimulation on mechanoreceptive and nociceptive afferent units from the rabbit pinna. Brain Research 378 , 21 to 27.
Barker, J.N., Mitra, R.S., Griffiths, C.E., Dixit, V.M.& Nicholoff, B.J. (1991). Keratinocytes as initiators of inflammation. Lancet 337 , 211 to 214.
Barrows, H.S., Tamblyn, R.N., (1980). Problem-based learning: an approach o medical education. Springer, New York.
Blottner, D.& Baumgarten, H.G. (1994). Neurotrophy and regeneration in vivo. Acta Anatomica 150 , 235 to 245.
Bogduk, N. (1993). The anatomy and physiology of nociception. In : Crosbie J.& McConnell, J.(Eds.) (1993). Key Issues in Musculoskeletal Physiotherapy (Ch.3). Oxford : Butterworth-Heinemann Ltd.
Butler, D.S., (1991). Mobilisation of the Nervous System . Churchill Livingstone : Melbourne.
Chatani, K., Kawakami, M., Weinstein, J.N., Meller, S.T.& Gebhart, G.F. (1995). Characterization of thermal hyperalgesia, c-fos expression, and alterations in neuropeptides after mechanical irritation of the dorsal root ganglion. Spine 20 (3), 277 to 290.
Coderre, T.J., Basbaum, A.I.& Levine, J.D. (1989). Neural control of vascular permeability : interactions between primary afferents, as cells, and sympathetic efferents. Journal of Neurophysiology 62 , 48-58
Coderre, T.J., Chan, A.K., Helms, C.& Levine, J.D. (1991). Increasing sympathetic nerve terminal-dependent plasma extravasation correlates with decreased arthritic joint injury in rats. Neuroscience 40 , 185 to 189.
Coghill, R.C., Mayer, D.J., Price, D.D., (1993). Spinal cord coding of pain : The role of spatial recruitment and discharge frequency in nociception. Pain , 53 , 295 to 309.
Cohen, M.L., (1995). The clinical challenge of secondary hyperalgesia. Moving in on Pain (Abstracts). Adelaide, Australia.
Cohen, M.L., Quintner, J.L., (1993). Fibromyalgia syndrome, a problem of tautology. Lancet , 342 , 906 to 909.
Collingride, G.L.& Singer, W. (1990). Excitatory amino acid receptors and synaptic plasticity. Trends in Pharmacological Science 11 , 290 to 296.
Cornefjord, M., Takahashi, K., Matsui, H., Olmarker, K., Holm, S.& Rydevik, B. (1992). Impairment of nutritional transport at double level cauda equina compression: An experimental study. Neuro Orthopedics 13 , 107 to 112.
Dickenson, A.H.& Sullivan, A.F. (1987). Evidence for a role of the NMDA receptor in the frequency dependent potentiation of deep rat dorsal horn nociceptive neurons following C-fibre stimulation. Neuropharmacology 26 , 1235 to 1238.
Donnerer, J., Amann, R. & Lembeck, F. (1991). Neurogenic and non-neurogenic inflammation in the rat paw following chemical sympathectomy. Neuroscience 45 (3), 761 to 765.
Enwemeka, C.S. & Spielholz, N.I. (1992). Modulation of tendon growth and regeneration by electrical fields and currents. In : Currier, D.P.& Nelson, R.M. (Eds.) (1992). Dynamics of Human Biologic Tissues (ch10). Philadelphia : F.A. Davis Company.
Ferrell, W.R., Wood, L. & Baxendale, R.H. (1988). The effect on acute joint inflammation on flexion reflex excitability in the decerebrate, low-spinal cat. Quarterly Journal Of Experimental Physiology 73 , 95 to 102.
Galea, M.P. & Darien-Smith, I. (1995). Voluntary movement and pain : Focussing on action rather than perception. Moving in on Pain . Adelaide.
Gallin, J.I., Goldstein, I.M. & Snyderman, R. (Eds.) (1992). Inflammation : Basic Principles and Clinical Correlates (2nd ed.). New York : Raven Press.
Gogas, K.R., Presley, R.W., Levine, J.D. & Basbaum, A.,I., (1991). The antinociceptive action of supraspinal opioids results from an increase in descending inhibitory control : Correlation of nociceptive behaviour and C-fos expression. Neuroscience 42 (3), 617 to 628.
Gracely, R.H., Lynch, S.L. & Bennett, G.,J. (1992). Painful neuropathy : Altered central processing, maintained dynamically by peripheral input. Pain 51 , 175 to 194.
Greenbag, P.E., Brown, M.D., Pallares, V.S., Tompkins, J.S. & Mann, N.,H., (1988). Epidural anesthesia for lumbar spine surgery. Journal of Spinal Disorders 1 , 139 to 143.
Gonzales, R., Goldyne, M.E., Taiwo, Y.O. & Levine, J.D. (1989). Production of hyperalgesic prostaglandins by sympathetic postganglionic neurons. Journal of Neurochemistry 53 , 1595-1598.
Gonzales, R., Sherbourne, C.D., Goldyne, M.E. & Levine, J.D., (1991). Noradrenaline-induced prostaglandin production by sympathetic postganglionic neurons is mediated by alpha-2-adrenergic receptors. Journal of Neurochemistry 57 , 1145 to 1150.
Green, P.G., Basbaum, A.I., Helms, C. & Levine, J.D. (1991). Purinergic regulation of bradykinin-induced plasma extravasation and adjuvant-induced arthritis in the rat. Proceedings of the National Academy of Science , U.S.A. 88 , 4162 to 4165.
Green, P.G., Luo, J., Heller, P.H. & Levine, J.D. (1992). Modulation of bradykinin-induced plasma extravasation in the rat knee joint by sympathetic co-transmitters. Neuroscience 52 , 451-458.
Groen, G.J., Balget, B. & Drukker, J. (1988). The innervation of the spinal dura mater: anatomy and clinical considerations. Acta Neurochirurgica 92 , 39 to 46.
Groenblad, M., Weinstein, J.N. & Santavirta, S. (1991). Immunohistochemical observation on spinal tissue innervation. Acta Orthopaedic Scandinavia 62 (6), 614 to 622.
Groenblad, M., Virri, J., Tolonen, J., Seitsalo, S., Kaeaepae, E., Kankare, J., Myllynen, P. & Karaharju, E. (1994). A controlled immunohistochemical study of inflammatory cells in disc herniation tissue. Spine 19 (24), 2744 to 2751.
Handwerker, H.O., Forster, C. & Kirchhoff, C., (1991). Discharge patterns of human C-fibres induced by itching and burning stimuli. Journal of Neurophysiology 66 , 307 to 317.
Harvey, G.K., Toyka, K.V. & Hartung, H.-P. (1994). Effects of mast cell degranulation on blood-nerve barrier permeability and nerve conduction in vivo. Journal of the Neurological Sciences 125 , 102-109.
Heller, P.H., Green, P.G., Tanner, K.D., Miao, F.J.-P. & Levine, J.D. (1994). Peripheral neural contributions to inflammation. Progress in Pain Management (1st ed.)(pp31 to 42). Seattle : IASP Press.
Hodges, P., Richardson, C., (1996). Inefficient muscular stabilisation of the lumbar spine associated with low back pain : A motor control evaluation of transverse abdominis. Spine , 21 , 2640 to 2650.
Hodges, P., Richardson, C., Jull, G., (1996). Evaluation of the relationship between laboratory and clinical tests of transverse abdominis function. Physiotherapy Research International , 1 , 30 to 40.
Hu, S. & Zhu, J. (1989). Sympathetic facilitation of sustained discharges of polymodal nociceptors. Pain 38 , 85 to 90.
Hsieh, J-C., Stahle-Backdahl, M., Hagermark, O., Stone-Elander, S., Rosenquist, G., Ingvar, M., (1995). Traumatic nociceptive pain activates the hypothalamus and the periaqueductal gray: a positron emission tomography study. Pain , 64 , 303 to 314.
Issekutz, T.B. (1992). Inhibition of lymphocyte endothelial adhesion and in vivo lymphocyte migration to cutaneous inflammation by TA-3, a new monoclonal antibody to rat LFA-1. Journal of Immunology 149 , 3392 to 3402.
Jaenig, W. (1985). Organization of the lumbar sympathetic outflow to skeletal muscle and skin of the cat hindlimb and tail. Review of Physiology, Biochemistry, Pharmacology 102 , 119 to 213.
Jaenig, W. & Koltzenberg, M. (1991). Receptive properties of pial afferents. Pain 45 , 77 to 85.
Jones, M., (1995). Clinical Reasoning and Pain. Manual Therapy , 1 , 17 to 24.
Jull, G., Zito, G., Trott, P., Potter, H., Shirley, D., Richardson, C., (1997). Inter-examiner reliability to detect painful upper cervical joint dysfunction. Australian Journal of Physiotherapy , 43 , 2, 125 to 129.
Kalso, E.A., Sullivan, A.F., McQuay, H.J., Dickenson, A.H., Roques, B.P., (1993). Cross-tolerance between Mu Opioid and alpha-2 Adrenergic receptors, but not between Mu and Delta receptors in the spinal cord of the rat. The Journal of Pharmacology and Experimental Therapeutics , 265 (2), 551 to 558.
Kambin, P., Abda, S. & Kurpici, F. (1980). Intradiskal pressure and volume recording : Evaluation of normal and abnormal intervertebral disks. Clinical Orthopedics 146 , 140.
Kawakami, M., Weinstein, J.N., Chatani, K.-I., Spratt, K.F., Meller, S.T. & Gebhart, G.F. (1994a). Experimental lumbar radiculopathy : Behavioural and histologic changes in a model of radicular pain after spinal nerve root irritation with chromic gut ligatures in the rat. Spine 19 (16), 1795 to 1802.
Kawakami, M., Weinstein, J.N., Spratt, K.F., Chatani, K-I., Traub, R.J., Meller, S.T. & Gebhart, G.F. (1994b). Experimental lumbar radiculopathy : Immunohistochemical and quantitative demonstrations of pain induced by lumbar nerve root irritation of the rat. Spine 19 (16), 1780-1794.
Kobayashi, S., Yoshizawa, H., Hachiya, Y., Ukai, T. & Morita, T. (1993). Vasogenic edema induced by compression injury to the spinal nerve root: Distribution of intravenously injected protein tracers and Gadolinium-Enhanced Magnetic Resonance Imaging. Spine 18 (11), 1410 to 1424.
Koltzenburg, M., Kees, S., Budweiser, S., Ochs, G. & Toyka, K.V. (1994). The properties of unmyelinated nociceptive afferents change in a painful chronic constriction neuropathy. In : Gebhart, G.F., Hammond, D.L. & Jensen, T. S. (Eds.) (1994). Proceedings of the 7th World Congress on Pain : Progress in Pain and Management (2nd ed.) (Ch.35). Seattle : IASP Press.
Korkala, O., Gronblad, M., Liesi, P. & Karaharju, E. (1985). Immunohistochemical demonstration of nociceptors in the ligamentous structures of the lumbar spine. Spine 10 , 156 to 157.
Kuslich, S.D., Ulstrom, C.L. & Michael, C.J. (1991). The tissue origin of low back pain and sciatica: A report of pain response to tissue stimulation during operation on the lumbar spine using local anaesthesia. Orthopedic Clinics North America 22 , 181 to 187.
Laird, J.M.A. & Bennett, G.J. (1993). An electrophysiological study of dorsal horn neurons in the spinal cord of rats with an experimental peripheral neuropathy. Journal of Neurophysiology 69 , 2072 to 2085.
Laird, J.M.A. & Cervero, F. (1990). Tonic descending infuences on receptive-field properties of nociceptive dorsal horn neurons in sacral spinal cord of rat. Journal of Neurophysiology 63 (5), 1022 to 1032.
LaMotte, R.H., Shane, C.N., Simone, D.A. & Tsai, E.F.P. (1991). Neurogenic hyperalgesia : Psychophysical studies of underlying mechanisms. Journal of Neurophysiology , 66 , 190 to 211.
Levine, J.D., Dardick, S.J., Roizen, M.F., Helms, C. & Basbaum, A.I. (1986). The contribution of sensory afferents and sympathetic efferents to joint injury in experimental arthritis. Journal of Neuroscience 6 , 3923 to 3929.
Levine, J.D., Goldstine, J., Mayes, M., Moskowitz, M.A. & Basbaum, A.I. (1986). The neurotoxic effect of gold sodium thiomalate on the peripheral nerves of the rat. Arthritis and Rheumatism 29 , 897 to 901.
Levine, J.D., Taiwo, Y.O., Collins, S.D. & Tam, J.K. (1986a). Noradrenaline hyperalgesia is mediated through interaction with sympathetic postganglionic neurone terminals rather than activation of primary afferent nociceptors. Nature 323 , 158 to 160.
Lotz, M., Vaughn, J.H. & Carson, D. (1988). Effect of neuropeptides on production of inflammatory cytokines by human monocytes. Science 241 , 1218 to 1221.
Lovick, T.A., (1991). Interactions between descending pathways from the dorsal and ventrolateral periaquaductal gray matter in the rat. In ; Depaulis, A., Bandler, R.,(eds). The Midbrain Periaqueductal Gray Matter . Plenium Press, New York, 101 to 120.
Lima, D., Esteves, F. & Coimbra, A., (1994). C- fos activation by noxious input of spinal neurons projecting to the nucleus of the tractus solitarius in the rat. In : Gebhart, G.F., Hammond, D.L. & Jensen, T.S. (Eds.) (1994). Proceedings of the 7th World Congress on Pain : Progress in Pain and Management (2nd ed.) (Ch.30). Seattle : IASP Press.
Liu, J., Roughley, P.J. & Mort, J.S. (1991). Identification of human intervertebral disc stromelysin and its involvement in matrix degeneration. Journal of Orthopedic Research 9 , 568 to 575.
Lundborg, G., (1988). Nerve Injury and Repair . Edinburgh : Churchill Livingstone
MacMillan, J., Schaffer, J.L. & Kambin, P. (1991). Routes and incidence of communication of lumbar discs with surrounding neural structures. Spine 16 , 2, 167 to171.
Magal, E.L. (1990). Gangliosides prevent ischemia-induced down-regulation of protein kinase C in fetal rat brain. Journal of Neurochemistry 55 , 2126-2131.
Maitland, G.D. (Ed.) (1986). Vertebral Manipulation (5th ed.). London : Butterworths.
Maitland, G.,D., (Ed) (1991). Peripheral Manipulation . London : Butterworths.
Mao, J., Price, D.D., Coghill, R.C., Mayer, D.J. & Hayes, R.L. (1992). Spatial patterns of spinal cord [14C]-2-deoxyglucose metabolic activity in a rat model of painful peripheral mononeuropathy. Pain 50 , 89 to 100.
Markowitz, S., Saito, K., Buzzi, M.G. & Moskowitz, M.A. (1989). The development of neurogenic plasma extravasation in the rat dura mater does not depend upon the degranulation of mast cells. Brain Research 477 , 157 to 165.
Matsui, H., Olmarker, K., Cornefjord, M., Takahashi, K. & Rydevik, B. (1992). Local electrophysiological stimulation in experimental double level cauda equina compression. Spine , 17 (9), 1075-1078.
Maixner, W., Gracely, R.H., Zuniga, J.R., Humphrey, C.B & Bloodworth, G.R. (1990). Cardiovascular and sensory responses to forearm ischemia and dynamic hand exercise. American Journal of Physiology 259 , R1156 to R1163.
McKenzie, R.C. & Sauder, D.N. (1990). The role of keratinocyte cytokines in inflammation and immunity. Journal of Investigative Dermatology 95 , S105 to 107.
Morton, C.R., Siegel, J., Xiao, H.-M., Zimmermann, M., (1997). Modulation of cutaneous nociceptor activity by electrical stimulation in the brain stem does not inhibit the nociceptive excitation of dorsal horn neurons. Pain , 71 , 65 to 70.
Morgan, M.M., Sohn, J.-H., Lohof, A.M. Ben-Eliyahu, S. & Liebeskind, J.C. (1989). Characterization of stimulation-produced analgesia from the nucleus tractus solitarius in the rat. Brain Research 486 , 175 to 180.
Nakagawa, I., Omote, K., Kitahata, L.M., Collins, J.G. & Murata, K. (1990). Serotonergic mediation of spinal analgesia and its interaction with noradrenergic systems. Anesthesiology 73 , 474 to 478.
Nishizuka, Y. (1989). The family of protein kinase C for signal transduction. JAMA 262 , 1826 to 1833.
Ochoa, J.L. & Yarnitsky, D. (1993). Mechanical hyperalgesia in neuropathic pain patients : Dynamic and static subtypes. Annals of Neurology 33 , 465 to 472.
Olmarker, K., Rydevik, B., Holm, S. & Bagge, U. (1989). Effects of experimental, graded compression on blood flow in spinal nerve roots. A vital microscope study on the porcine cauda equina. Journal of Orthopedic Research 7 , 817 to 823.
Olmarker, K., Rydevik, B. & Nordborg, C. (1993). Autologous Nucleus Pulposus induces neurophysiological and histological changes in porcine cauda equina nerve roots. Spine 18 (11), 1425 to 1432.
Petersen, N.,P., Vicenzino, B., Wright, A., (1993). The effects of a cervical mobilisation technique on sympathetic outflow to the upper limb in normal subjects. Physiotherapy Theory and Practice , 9 , 149 to 156.
O?Sullivan, P., Twomey, L., Allison, G., Sinclair, J., Miller, K., Knox, J., (1997). Altered patterns of abdominal muscle activation in patients with chronic low back pain. Australian Journal of Physiotherapy , 43 , 91 to 98.
Post, C., Minor, B.G., Davies, M. & Archer, T. (1986). Analgesia induced by 5-hydroxytryptamine-depletion in rats. Brain Research 363 , 18 to 27.
Price, D.D., Long, S. & Huitt, C. (1992). Sensory testing of pathophysiological mechanisms of pain in patients with reflex sympathetic dystrophy. Pain 49 , 163 to 173.
Price, D.D., Mao, J. & Mayer, D.J. (1994). Central neural mechanisms of normal and abnormal pain states. In Fields, H.L. & Liebeskind, J.C. (Eds.)(1994). Progress in Pain Research and Management (1st ed.)(pp61-84). Seattle : IASP Press.
Proudfit, H.K. (1992). The behavioural pharmacology of the noradrenergic descending system. In : Besson, J.M. & Guilbaud, G. (Eds.)(1992). Towards the use of Noradrenergic Agonists for the Treatment of Pain (1st ed.). Amsterdam : Elsevier Science Publishers B.V.
Quintner, J.L., Cohen, M.L., (1994). Referred pain of peripheral nerve origin : an alternative to the 'myofascial pain construct'. Clinical Journal of Pain , 10 , 243 to 251.
Raja, S.N., Meyer, R.A. & Campbell, J.N. (1988). Peripheral mechanisms of somatic pain. Anesthesiology 68 , 571 to 590.
Rees, H., Sluka, K.A., Westlund, K.N. & Willis, W.D. (1994). Do dorsal root reflexes augment peripheral inflammation ? NeuroReport 5 , 821 to 824.
Ren, K., Randich, A. & Gebhart, G.F. (1990). Modulation of spinal nociceptive transmission from nuclei tractus solitarii : A relay for effects of vagal afferent stimulation. Journal of Neurophysiology 63 (5), 971 to 986.
Roberts, W.J. & Elardo, S.M. (1985). Sympathetic activation of unmyelinated mechanoceptors in cat skin. Brain Research 339 , 123 to 125.
Rothwell, N.J. & Hopkins, S.J. (1995). Cytokines and the nervous system II : actions and mechanisms of action. Trends in Neuroscience 18 , 130 to 136.
Rydevik, B.L., Brown, M.D. & Lundborg, G. (1984). Pathoanatomy and pathophysiology of nerve root compression. Spine 9 , 7 to 15.
Rydevik, B.L., Myers, R.R. & Powell, H.C. (1989). Pressure increase in the dorsal root ganglion following mechanical compression: Closed compartment syndrome in nerve roots. Spine 14 (6), 574 to 576.
Sandkuehler, J., Eblien-Zajjur, A., Fu, Q.-G. & Forster, C. (1995). Differential effects of spinilization on discharge patterns and discharge rates of simultaneously recorded nociceptive and non-nociceptive spinal dorsal horn neurons. Pain 60 , 55-65.
Sanjue, H. & Jun, Z. (1989). Sympathetic facilitation of sustained discharges of polymodal nociceptors. Pain 38 , 85-90.
Schmidt, R.F. (1990). Experimental arthritis. Pain (supp) 5 , S215.
Schmidt, R.F., Schaible, H.-G., Messlinger, K., Heppelmann, B., Hanesch, U. & Pawlak, M. (1994). Silent and active nociceptors : Structure, functions, and clinical implications. In : Gebhart, G.F., Hammond, D.L. & Jensen, T. S. (Eds.) (1994). Proceedings of the 7th World Congress on Pain : Progress in Pain and Management (Ch.16) (2nd ed.). Seattle : IASP Press.
Segond von Banchet G, Boettger MK, Fischer N, Gajda M, Brauer R, Schaible H-G (2009). Experimental arthritis causes tumor necrosis factor-alpha dependent inflitration of macrophages into rat dorsal root ganglia which correlates with pain-related behavior. Pain, 145, 151 to 159
Shindo, K., Tsunoda, S-T.& Shiozawa, Z. (1994). Muscle spasm induced sympathetic reflex bursts on microneurography in a case with pontine demyelination. Clinical Autonomic Research 4 , 299 to 302.
Schomburg, E.,D. & Steffens, H. (1992). On the spinal motor function of nociceptive afferents and enkephalins. In : Jami, L., Pierrot-Deseilligny, E. & Zytnicki, D. (Eds.)(1992). Muscle Afferents and Spinal Control of Movement (pp.395-400)(1st ed.). Oxford : Pergamon Press.
Suval, W.D., Duran, W.N., Boric, M.P., Hobson, R.W., Berendsen, P.B. & Ritter, A.B. (1987). Microvascular transport and endothelial cell alterations preceding skeletal muscle damage in ischemia and reperfusion injury. The American Journal of Surgery 154 , 211 to 218.
Taiwo, Y.O. & Levine, J.D. (1989). Prostaglandin effects after elimination of indirect hyperalgesic mechanisms in the skin of the rat. Brain Research 492 , 397-399.
Takahashi, K., Olmarker, K., Holm, S., Porter, R.W. & Rydevik, B. (1993). Double level cauda equina compression. An experimental study with continuous monitoring of intraneural blood flow in the porcine cauda equina. Journal of Orthopedic Research 11 (1), 104 to 109.
Thompson, S.W.N. & Woolf, C.J. (1991). Primary afferent-evoked prolonged potentials in the spinal cord and their central summation : Role of the NMDA receptor. In : Bond, M.R., Charlton, J.E. & Woolf, C.J. (Eds). Proceedings of the VIth World Congress on Pain (pp291-298). Amsterdam : Elsevier
Tillman, L.J. & Cummings, G.S. (1992). Biologic mechanisms of connective tissue mutability. In : Currier, D.P. & Nelson, R.M. (Eds.) (1992). Dynamics of Human Biologic Tissues (pp1-44). Philadelphia : F.A. Davis Company.
Torebjoerk, E. (1994). Nociceptor dynamics in humans. In : Gebhart, G.F., Hammond, D.L. & Jensen, T. S. (Eds.) (1994). Proceedings of the 7th World Congress on Pain : Progress in Pain and Management (Ch.19) (2nd ed.). Seattle : IASP Press.
Troisier, O. & Cypel, D. (1986). Discography: An element of decision. Clinical Orthopedics 206 , 70.
Urban, L. & Randic, M. (1984). Slow excitatory transmission in rat dorsal horn : Possible mediation by peptides. Brain Research 290 , 336 to 341.
Vaccarino, F., Guidotti, A. & Costa, E. (1987). Ganglioside inhibition of glutamate-mediated protein kinase C translocation in primary cultures of cerebellar neurons. Proceedings of the National Academy of Science , U.S.A. 34 , 8707.
Wall, P.D. (1995). Treatment of pain. Moving in on Pain . Adelaide.
Wall, P.D. & Devor, M. (1983). Sensory afferent impulse originate from dorsal root ganglia as well as from the periphery in normal and nerve injured rats. Pain 17 , 321 to 339.
Wall, P.D. & Melzak, R. (1989) (Eds.). Textbook of Pain , (2nd. ed.). Edinburgh : Churchill Livingstone.
Weinstein, J.N., Claverie, W. & Gibson, S. (1988). The pain of discography. Spine 13 (12), 1344 to 1348.
Williams, T.J. & Hellewell, P.G. (1992). Endothelial cell biology. Adhesions molecules involved in the microvascular inflammatory response. American Review of Respiratory Disease 146 , S45 to 50.
Wiltse, L.L., Fonseca, A.S., Amster, J., Dimartino, P. & Ravessoud, F.A. (1993). Relationship of the dura, Hofmann's ligaments, Batson's plexus, and a fibrovascular membrane lying on the posterior surface of the vertebral bodies and attaching to the deep layer of the posterior longitudinal ligament: An anatomical, radiological, and clinical study. Spine 18 (8), 1030 to 1043.
Woolf, C.J. (1983). Evidence for a central component of post-injury pain hypersensitivity. Nature 306 , 686 to 688.
Woolf, C.J. (1984). Long-term alterations in the excitability of the flexion reflex produced by peripheral tissue injury in the chronic decerebrate rat. Pain 18 , 325 to 343.
Woolf, C.J. & McMahon, S.B. (1985). Injury-induced plasticity of the fexor reflex in chronic decerebrate rats. Neuroscience 16 , 395 to 404.
Woolf, C.J., Shortland, P. & Sivilotti, L.G. (1994). Sensitization of high mechanothreshold superficial dorsal horn and flexor motor neurones following chemosensitive primary afferent activation. Pain 58 , 141 to 155.
Woolf, C.J. & Swett, J.E. (1984). The cutaneous contribution to the hamstring flexor reflex in the rat : an electrophysiological and anatomical study. Brain Research 303 , 299 to 312.
Woolf, C.J. & Wall, P.D. (1986). Relative effectivenss of C primary afferent fibres in evoking a prolonged facilitation of the flexor reflex in the rat. The Journal of Neuroscience 6 , 1433 to 1442.
Wright, A., Vicenzino, B., (1995). Cervical mobilisation techniques, sympathetic nervous system effects and their relationship to analgesia. In : Schacklock, M.,O., (ed.). (1995). Moving in on Pain , Butterworth-Heineman, 164 to 173.
Xavier, A.V., Farrell, C.E., McDanal, J. & Kissin, I. (1990). Does antidromic activation of nociceptors play a role in sciatic radicular pain? Pain 40 , 77 to 79.
Zochodne, D.W. (1993). Epineural peptides : A role in neuropathic pain? Canadian Journal of Neurological Sciences 20 (1), 69 to 72.
Initially written June 1995, Last update : 16 October 2009
 What We do
What We do What We Treat
What We Treat