Eine Klinische, Neurophysiologische und Biomechanische Richtung in der Dosierung der Behandlung von Wirbelsaeule - Schulter Dysfunktion.
Martin Krause 1996
Einfuehrung
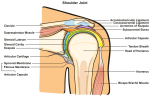
Schulter Dysfunktion kann verschiedene Urspruenge haben. Biomechanische Urspruenge der Dysfunktion deuten eine Instabilitaet an. Oefters wird der Subscapularis Muskel als Hauptfaktor in der Rehabilitation ange deutet. Nicht nur helfen solche einfachen Behandlungen nur wenigen Patienten, sondern es kann auch ein stoerender Faktor in dem Gedankenprozess des Therapeuten sein. Klinische Behandlungen und Neurophysiologische Erklaerungen von Schulter Dysfunktion gebrauchen vielfaeltige Dimensionen von Schmerz, Entzuendungsprozessen, und Kontrolle der Motorik um umfassende Erklaerungen zu bringen. Leider koennen solche vielfaeltigen Gedankensprozesse auch als Stoerfaktor wirken wenn die Organisationsstruktur des Wissens des Therapeuten nicht klar ist. Dieser Artikel wird von den organisierten Konzepten Geoffery Maitland's (1991) (Abb 1) und vom 'Klinisches Denken' (clinical reasoning) (Abb 2) ausgehen um die Bedeutung der Biomechanik (Abb 2-4) und Neurophysiologie (Abb.5) in dem Krankheitsbild zu erklaeren.
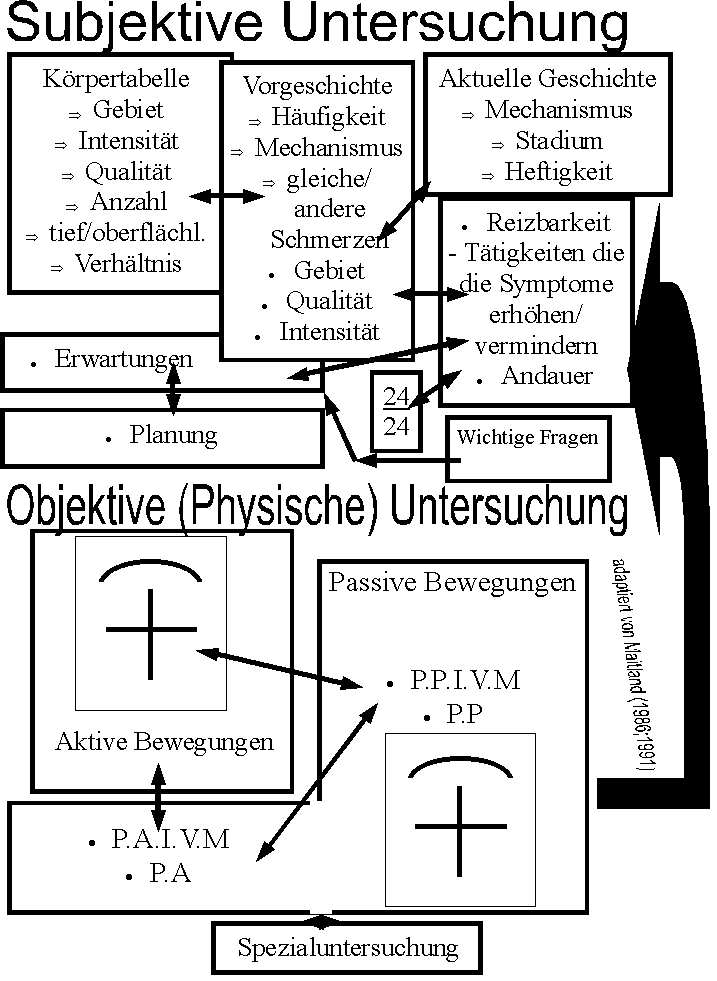
Abb 1 : Klinisches Denken und Kognitive Strukturierung des Maitlands Konzept
Neurogenische und Biomekanische Stabilitaet
Im Gegensatz zur Huefte kann die Schulter wenig Stabilitaet von den Ligumentaeren und vonder Knochenorientierung bekommen (Paavolainen et al 1984; Sarrafian 1983). Statt- dessen hat man postuliert dass es die Koordination der Muskulatur und des Bandapparates ist dass diese Stabilitaet erzeugt. Risse der Manschettenmuskulatur der Schulter hat man in drei kinematischen Dysfunktionen unterteilt. In dieser Unterteilung sind
'Stable Fulcrum Kinematics' verbunden mit Rissen des Supraspinatus Muskels (und Teile des Infraspinatus),
'Unstable Fulcrum Kinematics' verbunden mit grossen Rissen der oberen und hinteren Manschettenmuskulatur.
'Captured Fulkrum Kinematics' ist mit grossen Rissen der hinteren and voderen Manschettenmuskulatur verbunden und wirkt sich wie ein 'awning effect' unter den Akromion aus (Burkhart 1992).
Diese Studie deutet an dass wenn irgend ein Teil der Manschetten gerissen ist, dass es dann mindestens zu einem Schmerzhaftsbogen waehrend der Abduktion fuehren kann. Zum beispiel, kontraktion des Delta's erzeugt eine vertikale 'Shaerkraft' ohne Depression und Kompression der Manschettenmuskulatur (Baleman 1977). Es ist wahrscheinlich, dass wenn ein Komponent der Manschettenmuskulatur 'kompromiert' ist, dass dieses eine Auswirkung auf die restliche Manschettenmuskulatur, welches dass sich dann in 'Shaer' und Kompression der Blutgefaesse auswirkt, was zu einem Sauerstoffmangel fuehren kann (Poppen et al 1976; Post et al 1983).
Ein Schmerzhaftsbogen ist auch impliziert worden mit
' Verkuerzung der hinteren Kapsul und Schwaeche der Aussenrotatoren des Glenohumeralengelenkes
'Schwaeche der Innenrotatoren mit excessiver Aussenrotation des Glenohumeralengelenkes (Warner et al 1990).
Auch die lange Bicepsehne ist ein Teil der voderen Manschette und sollte in der Annamnesse betrachtet werden.
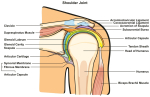
Fuer Abduktion braucht man unter anderem Aussenrotation des Humerus unterhalb des Akromions (Cailliet 1980; Sarrafian 1983; Schenkman und de Cartaya 1987). Weiterhin braucht man Rotation des Skapulas um eine effiziente Kontraktion ('line of pull') der Manschetten und Deltamuskulatur gewaehrleisten zu koennen (sehe Abb. 2) (Cailliet 1977). Ausserhalb der Klavikulabaender wird die Schulter von Muskeln des Skapulas 'aufgehaengt' (Baleman 1977) (sehe Abb.3). Die oberen Teile des Trapezius ziehen den Skapula hoch und nach innen um die Axen des Sternoklavikular und Akromioklavikulargelenkes (Cailliet 1980). Die mittleren Teile des Trapezius stabilisieren den Skapula und den unteren Teile ziehen den medialen Rand des Skapulas nach unten und dadurch erzeugen sie eine Rotation des Glenoideus nach oben. Durch diese Abduktion wechselt der Rotationsaxe von dem Glenohumeralengelenk, zum Sternoklavikularen- und Akromioklavikularengelenk. Um diese Stabilitaetsanderungen zu optimisieren braucht man auch die Kokontraktion des Serratus Anteriors, eine ekzentrische Kontraktion der Levator Skapulae, Rhomboidein, Pectoralis Minor, und Teres Major Muskulatur. Auch der Korakobrachialas und die Kurze-Biceps Muskel spielen eine Rolle waehrend der Abduktion. Dies bedeutet, dass die Muskulatur nicht als Agonist-Antagonist betrachtet werden sollte, sondern als eine koordinierte Mannschette (sehe Abb.2) (Kugler et al 1980; Schenkman et al 1987; Turvey et al 1978a, 1978b; 1981).
|
|
|
| Abb. 2 : Koordinierte Synergistische Stabilitaet durch Kontraktion des Schulterblatts und der Schultermuskulatur | |
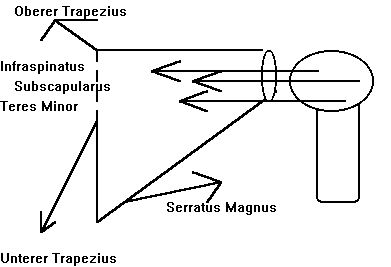
|
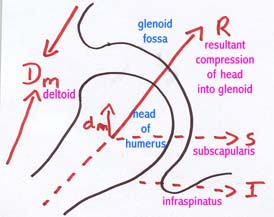
Abb. 3: Dynamische Stabilitaet des Schulterblatts |
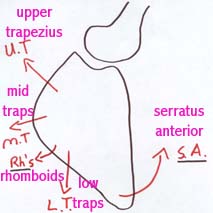
|
Abb. 4 : Unkoordinierte Schulterblatt Synergie |
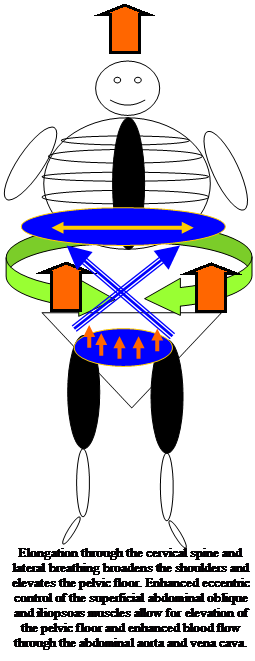
Auch das Zwerchfel ist von Bedeutung zur Schulter Problemen wobei die Luft im unteren Lungenbereich die Schulterblaettern hebt.
Objekt-Orientierte Kontrolle der Muskulatur
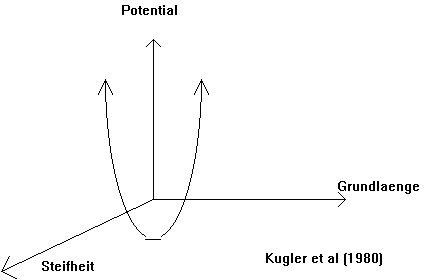
Eine Hypotese dieser koordinierten Stabilitaet ist, dass das Nervensystem wie ein 'Federungssystem' wirkt wo dynamische Stabilitaet durch 'Periodische Oscillierung' der gesamten elektrischen Aktivitaet der sensorisch-motorischen Neuronen darstellt (siehe Abb.5) (Kugler et al 1980; Segundo et al 1995; Jefferys et al 1996). Dies bedeutet, dass die Ausgangsphase jeder Bewegung der Schulter kritisch ist und dass diese in der funktionellen Behandlung von Schulterschmerzen betrachtet werden sollte. Was sich unterscheidet zwischen Bewegungen ist die Bedeutung dieser Bewegung zu der Funktion die erreicht werden moechte (Bernstein 1967; Glencross 1980; Kelso et al 1983; Turvey et al 1978; Reed 1982; Whiting 1980). Es ist wichtig zu verstehen, dass in diesem Konzept die Parameter 'Steifheit' und 'Grundlaenge' mit der gesamten elektrischen Aktivitaet im Nervensystem zusammen haengen (Kelso 1982).
|
Abb. 5 : 'Federungssytem' fuer dynamische Stabilitaet des Nervensystems. |
sehe : www.bodyblade.com
Klinisches Denken : Afferente-Efferente Imbalanz wegen Schmerzen und Entzuendungen
Die Bedeutung eines Kommandos von hoeheren Zentren des Nervensystems wird vom Grundstatus der elektrischen Aktivitaet in den Rueckenmarkneuronen beeinflusst. Der Grundstatus der elektrischen Aktivitaet wird von Afferenz und Efferenz gesteuert. Diese Afferenz kann durch Schmerzen erhoeht sein. Eine Reaktion einer erhoehten Afferenz resultiert in einer erhoehte Efferenz was eine erhoehte Spannung in der Muskulatur erzeugen kann (Gracely et al 1983; Woolf et al 1994; Woolf & Swett 1984). Das Maitland Konzept gebraucht Schmerz als Mittelpunkt der Behandlung von muskuloskeletischen Dysfunktionen (Abb.6). Um dieses Konzept zu verstehen sollte man 'klinisches Denken' und die Neurophysilogie der Biomechanik und der Schmerz in Betracht ziehen. Da der Organismus ueber das Nervensystem seine Existenz steuert, die richtigen Fragen stellen und Erklaerungen geben, koennten den Patienten seinen Kognitive Prozesse so steuern um die Optimale Erhohlung zu erreichen.
|
Abb 6: Klinisches Demken |
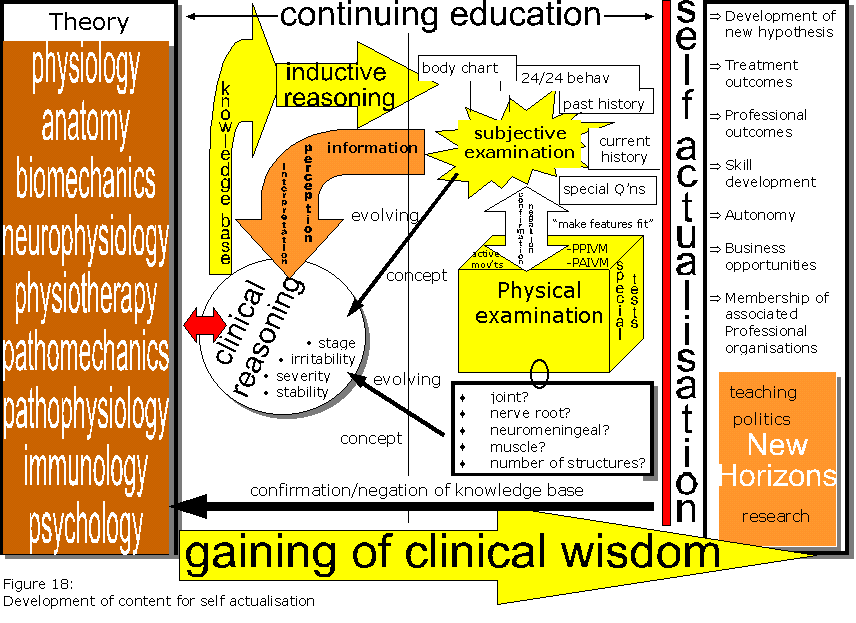
Moderne medizinische und paramedizinische Behandlung hat versucht die Krankheiten in Griff zu bekommen durch Diagnostizieren verschiedener Pathologien mit Benennungen wie z.B. Periarticulare Artrose, Omoartrose, usw. (Cohen & Quintner 1993; Quintner & Cohen 1994; Cohen 1995)
Fuer die Verbesserung der Geschicklichkeit der klinischen Begruendung koennte es fuer den Therapeuten wichtig sein die Pathophysiologie (die der Phenomenologie zugrunde liegt) besser zu verstehen.
Entzuendung
Die Reaktion des Gewebes nach einer Verletzung nennt man Entzuendung. Eine Entzuendung schliesst eine Serie von Vorgaengen in den Zellen (Enwemeka & Spielholz 1992; Schmidt et al 1994; Tillman & Cummings 1992). Eine Entzuendung kann der Grund fuer ein Oedem sein das als ?Irritant? auf Nervenrezeptoren, Nerven und deren Blutgefaesse wirken kann (Holthusen & Arndt 1996). Dieses kann Auswirkungen haben wo
- die Leitgeschwindichkeit der Nerven vermindert werden kann.
- erhoehte elektrische Aktivitaet der Nervenrezeptoren zu einer neurochemischen Reaktion im Rueckenmark fuehren kann.
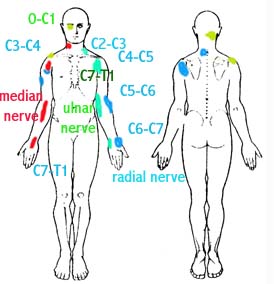
Weiterhin das heisst, dass Nerven mit gleicher Innervation wie das verletzte Gewebe entweder verminderte oder erhoehte motorische Aktivitaet haben koennte. Wenn es zu einer Verletzung der Wirbelsaeulestruktur kommt, dann koennte es zu einer Verspannung oder Entspannung von Muskeltonus in der Peripherie kommen wobei eine Instabilitaet und Bewegungsdysfunktion entsteht (z.B. Schmerzhaftsbogen). Ein Grund fuer verminderte Leitgeschwindichkeit kann durch einer Nervenkompromierung eines Oedems oder durch veraenderte neuronale Prozesse im Rueckenmark entstehen. Eine Kompromierung der Nerven kann nicht nur in der Wirbelsaeule entstehen (Abb.7), sondern auch in der Peripherie; z.B. koennte der Nervus Radialis zwischen Teres Major und Teres Minor kompromiert werden, der Nervus Supraskapularis kann im Fossa Supraskapularis kompromiert werden, das Plexus Brachialis kann unterhalb des Prozesus Korakoideus und des Pectoralis Minors kompromiert werden. Kompromierung unterschiedlicher Nerven koennte das 'Federungssytem' und somit die mechanische Stabilitaet des Glenohumeralengelenkes vermindern.
 |
|
Abb.7: Somatischer - Neurogenischer Innervations Bereiche |
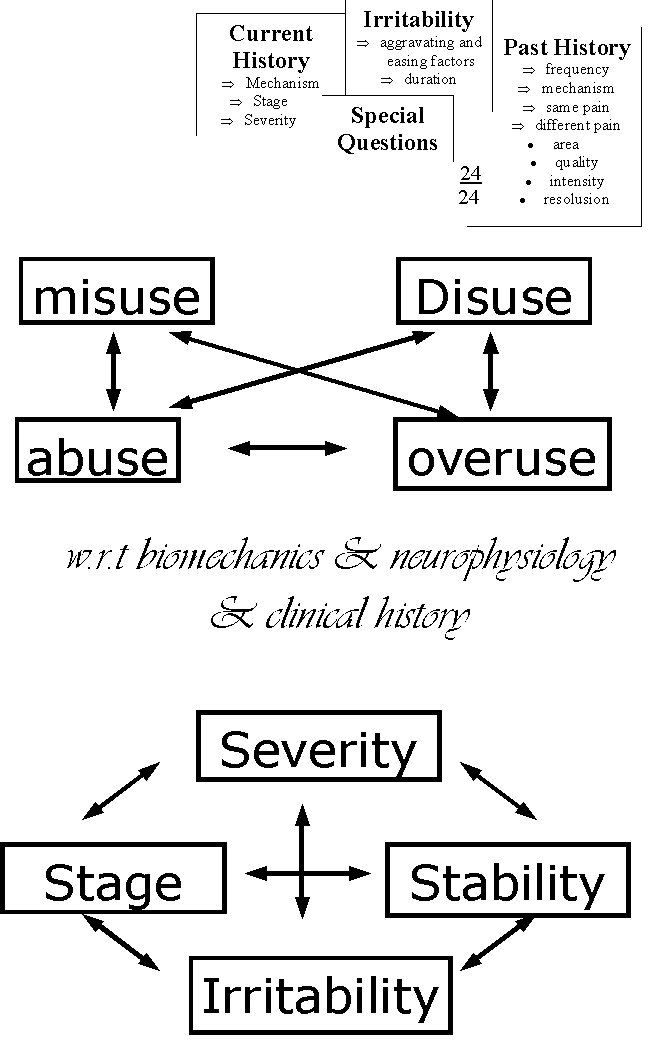
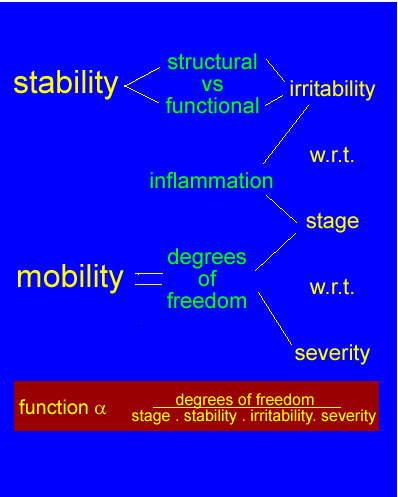
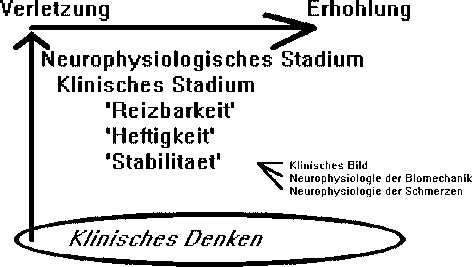
Klinische Begruendung einer solchen Hipotese entstehte wenn durch korrelation subjektiver und objektiver Untersuchungen ein Zusammenhang zwischen Wirbelsaule und Schulter gezeigt werden kann (Abb 1). Durch Behandlung der betroffenen Struktur sollte man eine Normalisierung der Anzeichen und Symptome zeigen koennen. Die Erwartung einer Behandlungsmethode sollte dann 'dem Stadium'(stage), 'der (Klinische und Neurophysiologischen [K&N]) Stabilitaet' (stability), 'der Heftigkeit' (severity) und 'der Reizbarkeit' (irritability) der Dysfunktion entsprechen (Maitland 1991) (Abb 8). Oefters kann man erwarten, dass z.B. neurologische Veraenderungen sich gleichzeitig normalisieren wenn eine Behandlungsmethode angewendet wird. Man kann auch eine Veaenderung des Schmerzhaftsbogen erwarten, wenn die richtige Behandlungsmethode fuer 'das Stadium'(stage), 'die (K&N) Stabilitaet'(stability), 'die Heftigkeit'(severity) und 'die Reizbahrkeit'(irritability) der Dysfunktion angewendet wird. Im Gegensatz kann man auch einer Verschlechterung der Anzeichen und Symptome erwarten wenn die Behandlungsmethode nicht dem klinischen Krankheitsbild entspricht. Das heisst auch, dass nicht nur die Behandlungsmethode genau sein muss sondern dass auch die Dosierung genau sein muss. Dosierung kann man mit Kraft, Richtung, Wiederholungen, Amplitude und Frequenz unterzeichnen : je mehr je hoeher die Dosierung. Man muss sich auch im Klaren sein, ob man das verletzte Gewebe mit erhoehtem mechanischem Reiz behandlen sollte oder ob man den mechanischen Reiz vermindern sollte; z.B. bei konstanten Schmerzen, grosser 'Heftigkeit' oder bei neurologischem Defizit in einem klinschen Bild einer Radikulopathie moechte man eher den mechanischen Reiz vermindern (Abb.9). Im Gegensatz, bei einem intermittierender Schmerz, kleiner 'Heftigkeit' moechte man eher den Schmerz provozieren um die Schwelle der Mechanorezeptoren zu erhoehen oder um die Modulierung der elektrischen Aktivitaet im Rueckenmark zu verbessern (Abb 10). Im diesem Konzept braucht eine minimale Dosierung um das Maximale verbesserung zu bekommen.
|
Abb. 8: Wichtige Aspekte des Klinisches Denkens Click on pic to view enlarged image |
|
Abb. 9: Verminderte Aktivierungsschwelle von Nervenrezptoren |
|
Abb. 10: Kortikale Elektrische Einflusse |

Neurophysiologische Modulierung von Schmerzen und Entzuendungen
Neurophysiologische Erklaerung der Wirkung von Tape (Klebverband) (wie bei McConnell), Nervenmobilisationen von Butler (1991), Gelenkmobilisationen von Maitland (1986;1991); Aktiv-Passiv Mobilisationen '(Mobilisation with Movement [MWM])' von Mulligan; 'Muskelenergie' von Babara Hungerford; Stabilisationsuebungen von Jull und 'posturelle Integrierung' von Shirley Sahrman haben eine aehnlichen Erklaerung. Diese Erklaerung entspricht das, dann wenn ein Organismus ueberleben moechte wird es versuchen die elektrische Aktivitaet im Nervensystem zu verringern wo es nicht erwuenscht ist, und zu optimieren wo es erwuenscht ist. Das bedeutet, dass dann wenn eine Entzuendung entsteht das Nervensystem versuchen wird diese konsequente elektrische Aktivitaet der Entzuendung zu verringern durch
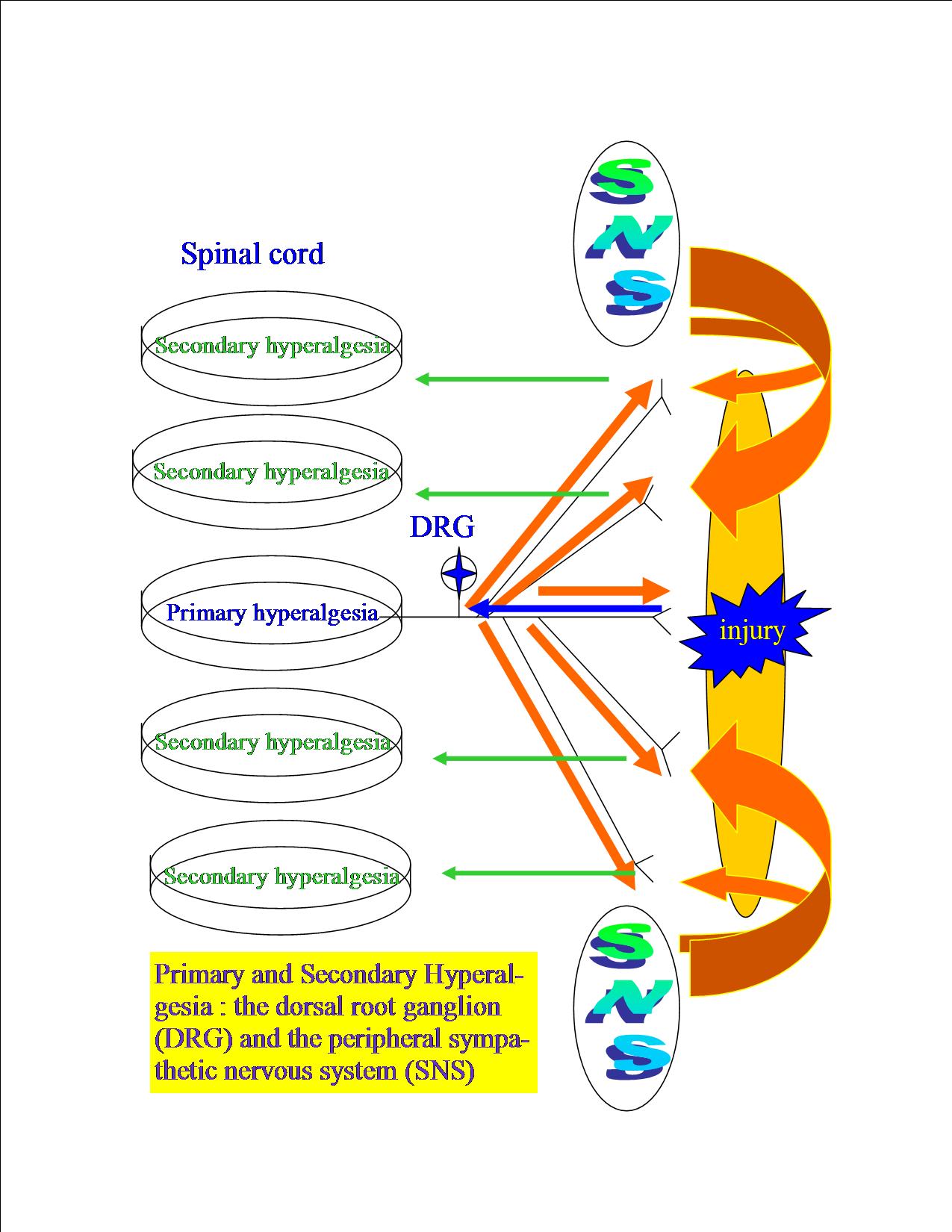
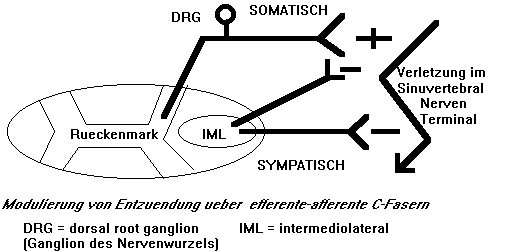
- Modulierung der Entzuendung durch das peripherale, symapthische und somatische Nervensystem (Abb.11),
- Modulierung der Nervenrezeptoren ueber das peripherale sympatische Nervensystem (SNS) (Abb.12),
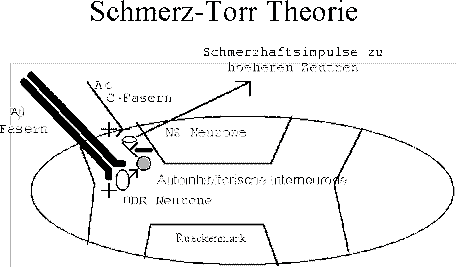
- Auto-Modulierung der Neuronen im Rueckenmark (Abb.13),
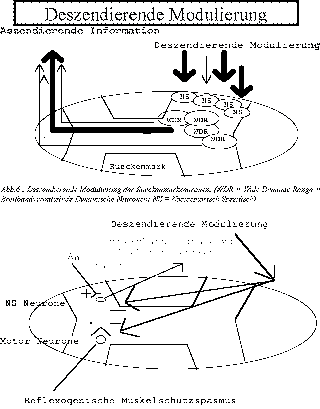
- deszendierende Modulierung der Rueckenmarkneuronen (Abb.14),
- deszendierende Modulierung des Schutzspasmus gewisser Muskulatur (Abb.14),
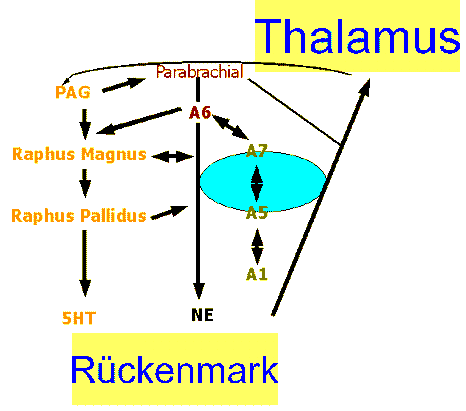
- Modulierung der Gehirn- und Rueckenmarkneuronen ueber dem Locus Coeruleus (Abb.15).
|
Abb.11 : Modulierung der Entzuendung durch das peripherale, sympathische und somatische Nervensystem |
|
Abb. 12 : Modulierung der Nervenrezeptoren ueber das peripherale sympatische Nervensystem (SNS) |
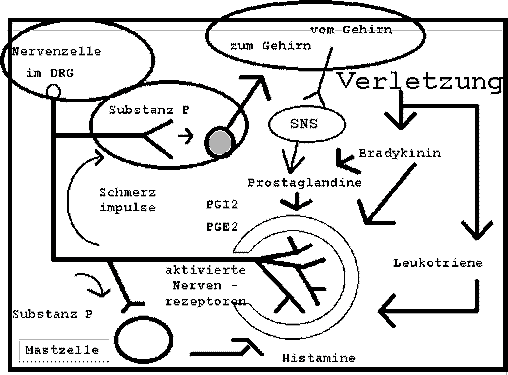
(adaptiert von : Groenblad et al 1991, p617; Jaenig 1985; Lotz et al 1988; Handwerker et al 1991; Levine et al 1986; Taiwo & Levine 1989).
|
Abb. 13 : Auto-Modulierung der Neuronen im Rueckenmark (NS = Schmerzvermittelnd Neurone; WDR Breitbandvermittelnde Dynamische Neurone) |
|
Abb. 14 : Deszendierende Modulierung des Muskelschutzspasmus |
|
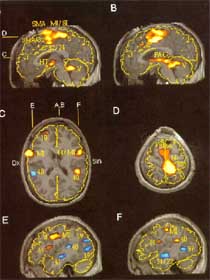
Abb.15 : Viele Kortikale Bereiche sind in das Modulierung des Schmerzens betaetigt (Hsieh, J-C., Stahle-Backdahl, M., Hagermark, O., Stone-Elander, S., Rosenquist, G., Ingvar, M., {1995}. Traumatic nociceptive pain activates the hypothalamus and the periaqueductal gray: a positron emission tomography study.) |
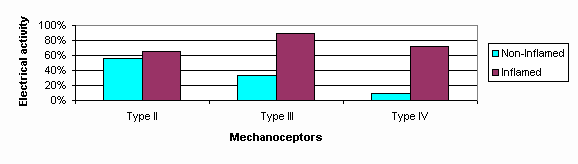
Eine neurogenische Entzuendung ist der Vorgang wobei freigelassene Neuropeptide der somatischen C-Fasern und sympatischen C-Fasern vaskulaere Membranveraenderungen bewirken welche zum Durchsickern von Proteinen des Gefaessystems in die umgebenden Gewebe fuehren (Ahmed et al 1991; Blottner & Baumgartner 1994; Donnerer et al 1991; Heller et al 1994; Kobayashi et al 1993; LaMotte et al 1991; Levine et al 1986; Markowitz et al 1989; Suval et al 1987; Zochodne 1993). Wenn die Entzuendung mechanischen Ursprung ist, dann erhoehen C-Fasern Rezeptoren ihre neurogenische Entzuendungsreaktion und erhoehen somit ihre Entladungsfrequenz mit Stimulation der sympatischen Nerventerminale (Levine et al 1986). Empfindlichkeitsfaehigkeit der peripheralen Nervenenden durch neurogenische und nicht-neurogenische Entzuendung koennte die Mechanorezeptorenschwelle verringern und dabei zum Schmerz leiten (siehe Abb.9).
Eine Reaktion auf Schmerzen ist eine erhoehte elektrische Aktivitaet in der Muskulatur die man im Tierexperiment auf den Versuch des Organismus zur Entlastung der elektrischen Aktivitaet der Mechanorezeptoren und Rueckenmarkneuronen zurueckfuehren kann (Abb.14) (Ferrell et al 1988; Woolf 1983; Woolf 1984; Woolf & McMahon 1985; Woolf & Wall 1986; Woolf et al 1994). Klinischer Beweis fuer Schutzspasmus kommt von Versuchen in Patienten mit cervikaler Radikulopathie wobei die Untersuchung eine erhoehte elektrische Aktivitaet in der innervierten Muskulatur gezeigt hat (Hall & Quintner 1996). Weiterhin wurde verminderte Beweglichkeit des Nervus Radialis im Epicondylitis des Ellenbogens auf Schmerzen und neurophysiologische Veraenderungen im Rueckenmark zurueckgefuehrt (Vicenzino, Collins, Wright 1996). Es ist wichtig zu wissen, dass man demonstrieren konnte, dass Manipulative Physiotherapie* der HWS diese Schmerzen im Ellenbogen vermindern konnte mit einer konsequenten Vermehrung der Beweglichkeit des Nervus Radialis (Vicenzino et al 1996).
- Manipulative Physiotherapie : das ganze Konzept der Manuellen Therapie gelehrt in Australien und nicht unbedingt eine Manipulation (Mobilisation mit Impulsen).
Extrapolation dieser Daten wuerde andeuten dass es Schutzspasmus verschiedenen Grades und in verschiedenen Gebieten gibt. Weil die Muskultur der Schulter und des Schulterblatt verschiedene Nerveninnervationen haben, ist es moeglich dass eine Dysfunktion der Wirbelsaulenstruktur zu einer Inkoordination von Schulter-Schulterblatt Muskulatur fuehren koennte (Abb.4). Eine Folge einer solchen Incoordination waere zu mindestens ein Schmerzhaftsbogen. Weiterhin, irgendeine konsequente Entzuendung der Schulterstrukturen koennte die Modulation der elektrischen Aktivitaet im Rueckenmark vermindern durch die (trophische) Einfluesse des innervierten Gewebes auf die Neuronen (Blottner & Baumgartner 1994). Dadurch koennte eine Hexenkreis zwischen Wirbelsaule und Schulter entstehen wobei beide Gebiete behandelt werden muessen.
Neurophysiologischer Beweis der Modulierung der entzuendeten Mechanorezeptoren und folglichem Heilungsprozess kommt von Studien des peripheralen sympatischen Nervensystems (Abb.11 und Abb.12). Die sympatischen postganglionischen Nerven entladen Entzuendungsvermittler, die die Plasmaextravasation, einschliesslich der Prostaglandine, erhoeht (Coderre et al 1989; Gonzales et al 1989; 1991; Green et al 1991). Ausserdem hat man gefunden, dass im Anwesen sympatischer postganglionischer neuronaler Modulierung, Plasmaextravasation durch Bradykinin erhoeht wurde (Green et al 1992). Die sympatischen, postganglionischen Neuronen geben auch Vermittler ab, die Plasmaextravasation verringern (Green et al 1991;1992). Ausser der Modulierung der Plasmaextravasation koennen diese Substanzen auch mit dem endothelialen Entspannungsfaktor (Greenberg et al 1991) und dem Blutplaettchenaktivierungsfaktor (Heller et al 1994) in den Blutgefaessen aufeinander einwirken. Der Mechanismus, wobei die symaptischen Nervenenden die Gewebeverletzung verringen, ist nicht geklaert. Es kann jedoch eine erhoehte Plasmaextravastion erwartet werden, wenn die Konzentration der Entzuendungssubstanzen durch die Facilitierung Lymph- und Venendraenage bewirkt wurde. Dadurch wird dann die Heilung beschleunigt (Heller et al 1994). Untersuchungen deuten an, das der Oedem dieses Entzuendungsprozesses nur im Zusammenhang mit Schmerzen entstehen kann (Holthusen & Arndt 1996). Wie beim Maitland Konzept ist eine Verringern der Schmerzen mit einer Resolution der Dysfunktion verbunden. Es ist wichtig zu verstehen dass bei intermittierenden (nicht-reizbaren) Schmerzen die Reproduzierung der Symptome manchmal wichtig ist um die Heilungsprozesse und die Normalisierung von Anzeichen und Symptomen zu beschleunigen (Maitland 1991).
Auto-Modulation elektrischer Aktivitaet der Rueckenmarkneuronen kann durch ein Oedem verringert werden (Abb.13). Eine Entzuendung kann der Grund fuer ein Oedem und Wirbel-Schulter Schmerzen sein (Barker et al 1991; Gallin et al 1992; Groenblad et al 1991; 1994b; Kawakami et al 1994a; 1994b; McKenzie & Saunder 1990; Rothwell & Hopkins 1995; Rydevik et al 1989; Williams & Hellewell 1992). Waehrend sich das Oedem vergroessert, koennen Kompression der intraneuralen Blutgefaesse und Nervengewebe vorzugsweise die Leitungsgeschwindigkeit der myelinischen Empfindungsnerven und der motorischen Nerven verringern und dabei wahrscheinlich die Leitungsgeschwindigkeit entlang der nicht-myelinischen Nervengewebe groesstenteils unbeeinflusst lassen (Cornefjord et al 1992; Kobayashi et al 1993; Matsui et al 1992; Olmarker et al 1989; Rydevik et al 1984; Takahashi et al 1993). Die verringerte Leitung entlang myelinischen Nervengewebe mit grossem Durchmesser entspricht einer konsequent verringerten Auto-Modulation der schmerzleitenden Neuronen im Rueckenmark (Dickenson & Sullivan 1987; Ochoa & Yarnitsky 1993; Price et al 1994). Als Resultat der Entzuendung ist zusaetzlich eine erhoehte elektrische Aktivitaet in den nicht-myelinischen Nerven zu erwarten. (Bennet & Xie 1988; Dubner 1991; Sugimoto et al 1990). Durch dosierte Manipulative Therapie wie z.B. Traktion, Rotation, Taping, oder spezifische Uebungen ist es moeglich die Leitgeschwindigkeit zu verbessern. Dieser wird bewirkt durch
- verbesserte Durchblutung (Selander, et al 1985).
- vermindertem Schutzspasmus (Abb.14)
- vermindertem Reiz auf Mechanorezeptoren
- verbesserte postganglionische, sympathische Modulierung der Mechanorezeptoren (Abb.11 und Abb.12)
- verbesserte postganglionische, sympathische Modulierung der Irrigation/Durchblutung
Leitung elektrischer Aktivitaet der Rueckenmarkneuronen als Folge eines Schmerzes kann auch von hoeheren Zentren stammen. Das zentrale, sympatische Nervensystem und spezifisch das Periaqueductale Grau spielt wahrsheinlich eine Rolle in Deszendierender Modulation der Schmerzleitung . Mindestens zwei moegliche Arten deszendierender Neuromodulation koennten angewendet werden (Bandler & Shipley 1994). Beide Arten deszendierender Modulation betrifft pontile, noradrenergische Projektion auf ventrale, dorsale und intermediolaterale (IML) Rueckenmarkneuronen (Proudfit 1992; Jaenig 1985; Morgan et al 1989; Nakagawa et al 1990; Post et al 1986; Ren et al 1990) Deszendierende Modulation wird als opiat- un abhaengige Analgesieart angegeben (Proudfit 1992) aber es gibt auch einen Beweis der die Meinung bestaetigt, dass m -opiat und a -2 noradrenalin Rezeptoren funktionell mit Schmerzmodulation verbunden sind (Kalso et al 1993). Die andere Art deszendierender Modulation wird als opiat-abhaengig angesehen (Bandler & Shipley 1994). Man glaubt, dass das noradrenalin System willkuehrlich motorische Aktivitaet vermehrt, wobei das opiat-abhaengige System Entspannung bewirkt (Bandler & Shipley 1994; Lovick 1991).
Klinische Untersuchungen haben beide diese Systeme als Grund gefunden fuer die Schnelle und andauernde Wirkung spezifischer Manipulativer Physiotherapie auf die Halswirbelsaeule (Wright & Vicenzino 1995). Eine Ueberpruefung der Akupunkturliteratur weisst darauf hin, dass der anfaengliche sympathoexcitatorische Effekt innerhalb 15 Sekunden von einer Sympathohemmung 20-45 Minuten spaeter gefolgt wird (Wright & Vicenzino 1995). Dieser Effekt konnte fuer 2 bis 40 Stunden andauern. Diese Anfangsergebnisse sind deshalb interessant weil es sehr wenige aufschlussreiche Studien gibt, die die neurophysiologischen und analgesischen Effekte der Manipulativen Physiotherapie untersuchen.
|
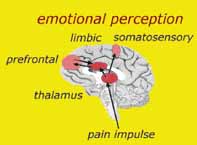
Abb. 16: Somatosensorische, Motorische und Limbische Bereiche werden bei der Untersuchung und Behandlung und Weideruntersuchung einbezogen |
Ausser den obengenannten deszendierenden Systemen der Schmerzmodulation werden noch andere Quellen in Betracht gezogen die von sensorimotorkortikalen Regionen stammen und mit Eingabe auf Rueckenmark- und den Hirnstammregionen zu tun haben (Wall 1995). Wall (1995) beschreibt dieses System als eine 'sensible Haltung' wobei Motoreingabe (Efferenz) eine starke Kontrolle ueber Sensorische (Afferente) Rueckenmarkvorgaenge ausuebt 'die sich mehr auf Aktion als auf Perzeption konzentriert'(p35; Galea & Darian-Smith 1995). Wall (1995) gebraucht Positive Emissions Tomograghie (PET) in Studien in Pateienten mit chronischen Schmerzen fuer den Beweis seiner Theorie, wohingegen Galea & Darian-Smith (1995) PET und anatomische Sektion von Primaten angewendet haben. Weiterer Beweis fuer deszendierende Modulation kommt von Gogas et al (1991) die fanden, dass supraspinale Opiate die Schmerzempfindung durch eine 64% Reduktion in oberflaechlicher Lamina-Hinterhornaktivitaet blockierten (gewoehnlich verbunden mit einer Ueberwiegenheit schmerzvermittelnder spezifischer Neuronen (NS) (Lima et al 1994)) und eine 85% Reduktion in Voderhornaktivitaet (gewoehnlich in Zusammenhang mit Motoraktivitaet) (Abb.14 und Abb.15) (Schomburg & Steffens 1991). Untersuchungen bestaetigten das Ergebnis, dass hoehere Zentren in einer anti-Schmerz vermittelnder Weise wirken koennen wo eine Ueberwiegenheit an Hemmung bei den NS Neuronen auftritt, wo aber Anregung und Hemmung mit gleicher Frequenz bei Breitenband Dynamischen Neuronen (WDR) auftritt (Sandkuehler et al 1995). Dies scheint eine besondere Denkweise zu sein wobei der kortikale Einfluss primaer zu einem sekuendaer ansteigenden schmerzvermittelenden Reiz ist. Es wird vorgeschlagen, dass das Nervensystem sehr spezifisch diskriminiert und dadurch auch die subjektive und physikalische Untersuchung, sowie die Dosierung der Behandlung sehr spezifisch sein muss fuer das 'Stadium', 'die (K&N) Stabilitaet', 'die Heftigkeit' und 'die Reizbarkeit' der Dysfunktion um die beste Normalisierung von Anzeichen und Symptomen zu erreichen (Abb.17).
|
Abb.17 : Korrelation zwischen Neurophysiologie (Theorie) und Klinischem Denken |
Auch die subjektive und physikalische Untersuchung kann ein wichtiger Teil der neurophysiologischen Modulierung der Schmerzen sein. Der Locus Coeruleus (LC) ist auch der Grund fuer Analgesia in Tieren und wahrscheinlich auch in Menschen (Fillenz 1990). Den LC hat man in der Modulierung von neuronaler Aktivitaet nicht nur im Rueckenmark sondern auch im kortikalem Bereich gefunden (Jefferys et al 1996; Fillenz 1991). Das LC braucht visuelle, akustische, und proprioceptorische Information um die Bedeutung der Stimulation der Behandlung und Untersuchung zu bewerten. Das bedeutet, dass eine Hypothese gemacht werden kann wobei eine Bewertung der Funktionsstoerungen und Behandlung LC's wird entscheiden wo es eine Dysfunktion gibt und ob die Behandlung diese elektrische Stoerung wieder normalisieren kann. Nicht nur der Therapeut/in muss wissen ob eine Bewegungsstoerung vorliegt, sondern der Patient muss es auch wissen (Abb. 18). Weiter muesste sowohl der Therapeut/in als auch der Patient eine klare Vorstellung haben wo die Ziele der Behandlung liegen. Es ist nicht genug zu glauben, dass die Schulterschmerzen von der Wirbelsaule kommen, oder dass Wirbelsaeulenschmerzen von der Schulter kommen, sondern man muss es auch zeigen koennen waehrend der Annamnese und waehrend der Behandlung. Wenn man genau weiss wo die Ziele liegen dann wird man auch wissen wann eine Behandlung wegen mangelder Fortschritte geaendert werden muss. Glauben ist nicht Wissen.
|
Abb. 18: Deszendierende Modulation ueber den Thalamus und Hirnstamm |
Durch eine Bewertung des 'Stadiums' und 'der Reizbarkeit' (Maitland 1991) der Dysfunktion sollte man feststellen koennen wie weit man mit der physikalische Untersuchung gehen moechte. Als Folge dieser beiden Untersuchungen kann man versuchen die genaue Dosierung der Behandlung der Dysfunktion anzupassen (Maitland 1991). Somit ist das Risiko verringert, dass sich die Symptome des Patienten durch die Behandlung verschlechtern. Faktoren die man in der Dosierung von Uebungen und Mobilisation in Betracht ziehen sollte sind :
- Konstant oder Intermittierend
- Belasten oder Entlasten
- Frequenz der Wiederholungen
- Wie viele Wiederholungen
- Dauer der Wiederholungen
- Kraftanwendung
- Kombinierte Bewegungen zum Belasten oder Entlasten
- Richtung der Anwendung
- Eine oder mehrere Strukturen belasten
Als facet; wenn bei einer Dysfunktion neurologische Veraendurungen in einer klaren radikularen Distribution vorliegen, und/oder wenn es in einem akuten oder verschlimmernten 'Stadium' ist, und/oder 'Reizbar' ist, dann gebraucht man eher Entlastungsmethoden die konstant (nicht-intermittierend) sind, mit wenig Kraft, wenigen Wiederhohlungen, wenig Dauer, kombinierte Bewegungen um zu Entlasten, tape (Klebverband) zum entlasten, Waerme/Kaelte und Elektroterapie. Wenn eine Dysfunktion im Heilungsstadium ist wobei es nicht 'Reizbar' ist, kann man die Dosierung vermehren um die Belastbahrkeit der Struktur bis zu 100% der zu erwarteten Resolution der Dysfunktion zu erhoehen. Man muss sich klar sein, ob man eine Struktur entlasten oder belasten moechte und die Antwort dieser Frage sollte im Zusammenhang mit Biomechanik, Neurophysiologie und klinischer Annamnese stehen.
|
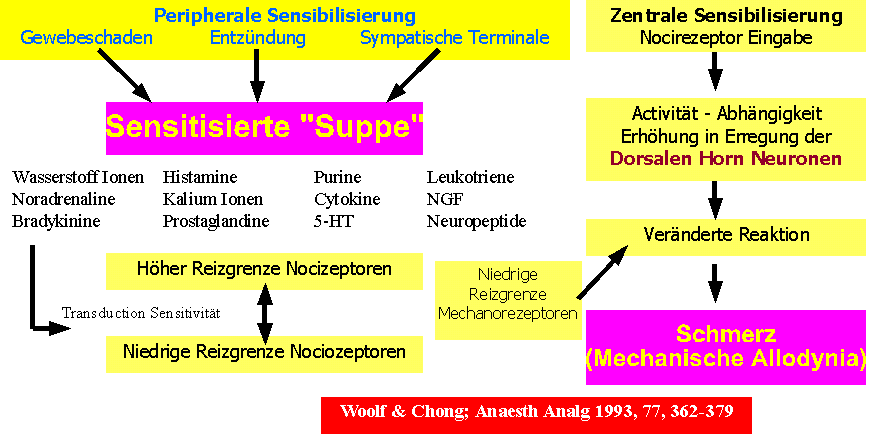
Abb 19: Eventuel kann eine gute und fruehe Behandlung der "Sensitisierte Suppe" mechanischen allodynia und chronischer Schmerz vermeiden? |
Sehe : Uebungen in Klinisches Denken (Deutsch)
Lendenwirbel Dysfunktion und Klinisches Denken
Geschrieben von Martin Krause hauptsaechtlich in 1996 und in 1986.
References
Ahmed, M., Bjurholm, A., Kreicbergs, A.& Schultzberg, M. (1993). Sensory and autonomic innervation of the facet joint in the rat lumbar spine. Spine 18 (14), 2121 to 2126.
Baleman, J.E., (1977). The Shoulder and Neck (Chapter 3). W.B. Saunders. Philadelphia.
Bandler, R., Shipley, M.T., (1994). Columnar organization in the midbrain periaqueductal gray : modules for emotional expression? TINS , 17 , 9, 379 to 389.
Barker, J.N., Mitra, R.S., Griffiths, C.E., Dixit, V.M.& Nicholoff, B.,J. (1991). Keratinocytes as initiators of inflammation. Lancet 337 , 211 to 214.
Bennett, G.,J., Xie, Y.,K., (1988). A peripheral mononeuropathy in the rat produces disorders of pain sensation like those seen in man. Pain , 33 , 87 to 107.
Bernstein, N., (1967). The Coordination and Regulation of Blood . Pergamon. New York.
Blottner, D.& Baumgarten, H.G. (1994). Neurotrophy and regeneration in vivo. Acta Anatomica 150 , 235 to 245.
Burkhart, S.,S. (1992). Fluoroscopic comparison of kinematic patterns in massive rotator cuff tears : a suspension bridge model. Clinical Orthopaedics and Related Research , 284 , 144 to 152.
Butler, D.S., (1991). Mobilisation of the Nervous System . Churchill Livingston. Melbourne
Cailliet, R., (1980). The Shoulder in Hemiplegia . E.A. Davis Co. Philadelphia.
Coderre, T.J., Chan, A.K., Helms, C.& Levine, J.D. (1991). Increasing sympathetic nerve terminal-dependent plasma extravasation correlates with decreased arthritic joint injury in rats. Neuroscience 40 , 185 to 189.
Cohen, M.,L., (1995). The clinical challenge of secondary hyperalgesia. Moving in on Pain (Abstracts). Adelaide, Australia.
Cohen, M.,L., Quintner, J.,L., (1993). Fibromyalgia syndrome, a problem of tautology. Lancet , 342 , 906 to 909.
Cornefjord, M., Takahashi, K., Matsui, H., Olmarker, K., Holm, S.& Rydevik, B. (1992). Impairment of nutritional transport at double level cauda equina compression: An experimental study. Neuro Orthopedics 13 , 107 to 112.
Dickenson, A.H.& Sullivan, A.F. (1987). Evidence for a role of the NMDA receptor in the frequency dependent potentiation of deep rat dorsal horn nociceptive neurons following C-fibre stimulation. Neuropharmacology 26 , 1235 to 1238.
Donnerer, J., Amann, R. & Lembeck, F. (1991). Neurogenic and non-neurogenic inflammation in the rat paw following chemical sympathectomy. Neuroscience 45 (3), 761 to 765.
Dubner, R., (1991). Neuronal plasticity and pain following peripheral tissue inflammation or nerve injury. Cited in : Bond, M.,R., Charlton, J.,E., Woolf, C.,J., (Eds) (1991). Proceedings of the VIth World Congress on Pain . Pain Research and Clinical Management (5th ed)(pp263 to 276). Amsterdam : Elsevier.
Enwemeka, C.S. & Spielholz, N.I. (1992). Modulation of tendon growth and regeneration by electrical fields and currents. In : Currier, D.P.& Nelson, R.M. (Eds.) (1992). Dynamics of Human Biologic Tissues (ch10). Philadelphia : F.A. Davis Company.
Ferrell, W.R., Wood, L. & Baxendale, R.H. (1988). The effect on acute joint inflammation on flexion reflex excitability in the decerebrate, low-spinal cat. Quarterly Journal Of Experimental Physiology 73 , 95 to 102.
Fillenz, M., (1990). Noradrenergic neurons . Cambridge University Press. Cambridge.
Galea, M.P. & Darien-Smith, I. (1995). Voluntary movement and pain : Focussing on action rather than perception. In : Shackock, M., (Ed). Moving in on Pain . Adelaide.
Gallin, J.I., Goldstein, I.M. & Snyderman, R. (Eds.) (1992). Inflammation : Basic Principles and Clinical Correlates (2nd ed.). New York : Raven Press.
Glencross, D.J., (1980). Levels and strategies of response organizations. In : Stelmach, G.e., Requin, J., (Eds). Tutorials in Motor Behavior . Nth. Holland Pub. Co..
Gogas, K.R., Presley, R.W., Levine, J.D. & Basbaum, A.,I., (1991). The antinociceptive action of supraspinal opioids results from an increase in descending inhibitory control : Correlation of nociceptive behaviour and C-fos expression. Neuroscience 42 (3), 617-628.
Gracely, R.H., Lynch, S.L., Bennett, G.J., (1992). Painful neuropathy : Altered central processing maintained dynamically by peripheral input. Pain , 51 , 175 to 194.
Gonzales, R., Goldyne, M.E., Taiwo, Y.O. & Levine, J.,D. (1989). Production of hyperalgesic prostaglandins by sympathetic postganglionic neurons. Journal of Neurochemistry 53 , 1595 to 1598.
Gonzales, R., Sherbourne, C.D., Goldyne, M.E. & Levine, J.,D., (1991). Noradrenaline-induced prostaglandin production by sympathetic postganglionic neurons is mediated by alpha-2-adrenergic receptors. Journal of Neurochemistry 57 , 1145 to 1150.
Green, P.G., Basbaum, A.I., Helms, C. & Levine, J.,D. (1991). Purinergic regulation of bradykinin-induced plasma extravasation and adjuvant-induced arthritis in the rat. Proceedings of the National Academy of Science , U.S.A. 88 , 4162 to 4165.
Green, P.G., Luo, J., Heller, P.H. & Levine, J.,D. (1992). Modulation of bradykinin-induced plasma extravasation in the rat knee joint by sympathetic co-transmitters. Neuroscience 52 , 451 to 458.
Greenberg, S.,S., Peevy, K., Tanaka, T.,P., (1991). Endothelium-derived and intraneuronal nitric oxide-dependent inhibition of norepinephrine efflux from sympathetic nerves by bradykinin. American Journal of Hypertension , 5 , 464-467. Cited in Heller, P.,H., Green, P.,G., Tanner, K.,D., Miao, F.,J.-P., Levine, J.,D., (1994). Peripheral neural contributions to inflammation. In Fields, H.,L., Liebeskind, J.,C., (Eds.). Progress in Pain Management (1st ed.)(31 to 42). Seattle : IASP Press.
Groenblad, M., Weinstein, J.N. & Santavirta, S. (1991). Immunohistochemical observation on spinal tissue innervation. Acta Orthopaedic Scandinavia 62 (6), 614 to 622.
Hall, T., Quintner, J., (1996). Response to mechanical stimulation of the upper limb in painful cervical radiculopathy. Australian Journal of Physiotherapy , 42 , 4, 277 to 286.
Handwerker, H.O., Forster, C. & Kirchhoff, C., (1991). Discharge patterns of human C-fibres induced by itching and burning stimuli. Journal of Neurophysiology 66 , 307 to 317.
Heller, P.H., Green, P.G., Tanner, K.D., Miao, F.J.-P. & Levine, J.D. (1994). Peripheral neural contributions to inflammation. Progress in Pain Management (1st ed.)(pp31-42). Seattle : IASP Press.
Holthusen, H., Arndt, J.O., (1996). The role of pain from veins for the formation of perivenous edema in humans. Pain , 68 , 395 to 400.
Jaenig, W. (1985). Organization of the lumbar sympathetic outflow to skeletal muscle and skin of the cat hindlimb and tail. Review of Physiology, Biochemistry, Pharmacology 102 , 119 to 213.
Jefferys, J.G.R., Traub, R.D., Whittington, M.A., (1996). Neuronal networks for induced ?40Hz?rhythms. TINS , 19 , 5, 202 to 208.
Kalso, E.,A., Sullivan, A.,F., McQuay, H.,J., Dickenson, A.,H., Roques, B.,P., (1993). Cross-tolerance between Mu Opioid and alpha-2 Adrenergic receptors, but not between Mu and Delta receptors in the spinal cord of the rat. The Journal of Pharmacology and Experimental Therapeutics , 265 (2), 551 to 558.
Kelso, J.A.S., (1982). Part 1 : Perspectives and issues in motor behavior. In ; Kelso, J.A.S (Ed). The Process Approach to Understanding Human Motor Behavior : an Introduction . Erlbaum. New York.
Kelso, J.A.S., Putman, C.A., Goodman, D., (1983). On the space-time structure of human interlimb co-ordination. Quarterly Journal of Experimental Psychology , 35A , 347 to 375.
Kobayashi, S., Yoshizawa, H., Hachiya, Y., Ukai, T. & Morita, T. (1993). Vasogenic edema induced by compression injury to the spinal nerve root: Distribution of intravenously injected protein tracers and Gadolinium-Enhanced Magnetic Resonance Imaging. Spine 18 (11), 1410 to 1424.
Kuglar, P.N., Kelso, J.A.S., Turvey, M.T., (1980). On the concept of coordinated structures as dissipative structures. I : Theoretical lines of converged tutorials in motor behavior. p3-45. II : Emperical lines of convergence. pp 49-70. In : Stelmach, G.E., Requin, J., (Eds). Tutorials in Motor Behavior . Nth Holland Publ. Co. pp3 to 45.
LaMotte, R.H., Shane, C.N., Simone, D.A. & Tsai, E.F.P. (1991). Neurogenic hyperalgesia : Psychophysical studies of underlying mechanisms. Journal of Neurophysiology , 66 , 190 to 211
Levine, J.D., Taiwo, Y.O., Collins, S.D. & Tam, J.K. (1986a). Noradrenaline hyperalgesia is mediated through interaction with sympathetic postganglionic neurone terminals rather than activation of primary afferent nociceptors. Nature 323 , 158 to 160.
Levine, J.D., Dardick, S.J., Roizen, M.F., Helms, C. & Basbaum, A.I. (1986). The contribution of sensory afferents and sympathetic efferents to joint injury in experimental arthritis. Journal of Neuroscience 6 , 3923 to 3929.
Lima, D., Esteves, F. & Coimbra, A., (1994). C- fos activation by noxious input of spinal neurons projecting to the nucleus of the tractus solitarius in the rat. In : Gebhart, G.F., Hammond, D.L. & Jensen, T.S. (Eds.) (1994). Proceedings of the 7th World Congress on Pain : Progress in Pain and Management (2nd ed.) (Ch.30). Seattle : IASP Press.
Lovik, T., (1991). Interactions between descending pathways from the dorsal and ventrolateral periaqueductal gray matter in the rat. In: Depaulis, A., Bandler, R., (Eds) (1990). The Midbrain Periaqueductal Gray Matter , 213 , 101 to 120. Plenium Press. New York.
Lotz, M., Vaughn, J.H. & Carson, D. (1988). Effect of neuropeptides on production of inflammatory cytokines by human monocytes. Science 241 , 1218 to 1221.
Maitland, G.D. (Ed.) (1986). Vertebral Manipulation (5th ed.). London : Butterworths.
Maitland, G.,D., (Ed) (1991). Peripheral Manipulation . London : Butterworths.
Matsui, H., Olmarker, K., Cornefjord, M., Takahashi, K. & Rydevik, B. (1992). Local electrophysiological stimulation in experimental double level cauda equina compression. Spine , 17 (9), 1075 to 1078.
Markowitz, S., Saito, K., Buzzi, M.G. & Moskowitz, M.A. (1989). The development of neurogenic plasma extravasation in the rat dura mater does not depend upon the degranulation of mast cells. Brain Research 477 , 157 to 165.
McKenzie, R.C. & Sauder, D.N. (1990). The role of keratinocyte cytokines in inflammation and immunity. Journal of Investigative Dermatology 95 , S105 to 107.
Morgan, M.M., Sohn, J.-H., Lohof, A.M. Ben-Eliyahu, S. & Liebeskind, J.C. (1989). Characterization of stimulation-produced analgesia from the nucleus tractus solitarius in the rat. Brain Research 486 , 175-180.
Nakagawa, I., Omote, K., Kitahata, L.M., Collins, J.G. & Murata, K. (1990). Serotonergic mediation of spinal analgesia and its interaction with noradrenergic systems. Anesthesiology 73 , 474 to 478.
Ochoa, J.L. & Yarnitsky, D. (1993). Mechanical hyperalgesia in neuropathic pain patients : Dynamic and static subtypes. Annals of Neurology 33 , 465 to 472.
Olmarker, K., Rydevik, B., Holm, S. & Bagge, U. (1989). Effects of experimental, graded compression on blood flow in spinal nerve roots. A vital microscope study on the porcine cauda equina. Journal of Orthopedic Research 7 , 817 to 823.
Paavolainen, P., Bjoerkenheim, J.M., Ahovuo, J., Slaetis, P., (1984). Recurrent anterior dislocation of the shoulder. Acta Orthopaedica , 55 , 556 to 560.
Poppen, N.K., Walker, P.S., (1976). Normal and abnormal motion of the shoulder. The Journal of Joint and Bone Surgery , 58A , 2, 195 to 200.
Post, M., Silver, R., Singh, M., (1983). Rotator cuff tear. Clinical Orthopedics of North America , 173 , 3, 78 to 91.
Proudfit, H.K., (1992). The behavioural pharmacology of the noradrenergic descending system. In : Besson, J.M., Guilbaud, G., (Eds) (1992). Toward the use of Noradrenergic Agonists for the Treatment of Pain. 119 to 136. Excerpta Medica. Amsterdam.
Price, D.D., Mao, J. & Mayer, D.J. (1994). Central neural mechanisms of normal and abnormal pain states. In Fields, H.L. & Liebeskind, J.C. (Eds.)(1994). Progress in Pain Research and Management (1st ed.)(pp61 to 84). Seattle : IASP Press.
Quintner, J.,L., Cohen, M.,L., (1994). Referred pain of peripheral nerve origin : an alternative to the 'myofascial pain construct'. Clinical Journal of Pain , 10 , 243 to251.
Reed, E., (1982). An outline of a theory of action systems. Journal of Motor Behavior , 14 , 2, 98 to 134.
Ren, K., Randich, A. & Gebhart, G.F. (1990). Modulation of spinal nociceptive transmission from nuclei tractus solitarii : A relay for effects of vagal afferent stimulation. Journal of Neurophysiology 63 (5), 971 to 986.
Rothwell, N.J. & Hopkins, S.J. (1995). Cytokines and the nervous system II : actions and mechanisms of action. Trends in Neuroscience 18 , 130 to 136.
Rydevik, B.L., Brown, M.D. & Lundborg, G. (1984). Pathoanatomy and pathophysiology of nerve root compression. Spine 9 , 7 to 15.
Rydevik, B.L., Myers, R.R. & Powell, H.C. (1989). Pressure increase in the dorsal root ganglion following mechanical compression: Closed compartment syndrome in nerve roots. Spine 14 (6), 574 to 576.
Sandkuehler, J., Eblien-Zajjur, A., Fu, Q.-G. & Forster, C. (1995). Differential effects of spinilization on discharge patterns and discharge rates of simultaneously recorded nociceptive and non-nociceptive spinal dorsal horn neurons. Pain 60 , 55 to 65.
Sarrafian, S.K., (1983). Gross and functional anatomy of the shoulder. Clinical Orthopedics and Related Research , 173 , 3, 11 to 19.
Schomburg, E.,D. & Steffens, H. (1992). On the spinal motor function of nociceptive afferents and enkephalins. In : Jami, L., Pierrot-Deseilligny, E. & Zytnicki, D. (Eds.)(1992). Muscle Afferents and Spinal Control of Movement (pp.395-400)(1st ed.). Oxford : Pergamon Press.
Schenkman, M., de Cartaya, V.R., (1987). Kinesiology of the shoulder complex. The Journal of Orthopedic Sports Physical Therapy , 8 , 9, 438 to 450.
Schmidt, R.F., Schaible, H.-G., Messlinger, K., Heppelmann, B., Hanesch, U. & Pawlak, M. (1994). Silent and active nociceptors : Structure, functions, and clinical implications. In : Gebhart, G.F., Hammond, D.L. & Jensen, T. S. (Eds.) (1994). Proceedings of the 7th World Congress on Pain : Progress in Pain and Management (Ch.16) (2nd ed.). Seattle : IASP Press .
Segundo, J.P., Vibert, J.F., Stiber, M., Hanneton, S., (1995). Periodically modulation inhibition and its postsynaptic consequences - I. General features. Inluence of modulation frequency. Neuroscience , 68 , 3, 657 to 692.
Selander, D., Mansson, L.,G., Karlsson, L., Svanvik, J., (1985). Adrenergic vasoconstriction in peripheral nerves of the rabbit. Anesthesiology , 62 , 6 to 10.
Sugimoto, T., Bennett, G.,J., Kajander, K.,C., (1990). Transsynaptic degeneration in the superficial dorsal horn after sciatic nerve injury : Effects of chronic constriction injury, transection, and strychnine. Pain , 42 , 205 to 213.
Suval, W.D., Duran, W.N., Boric, M.P., Hobson, R.W., Berendsen, P.B. & Ritter, A.B. (1987). Microvascular transport and endothelial cell alterations preceding skeletal muscle damage in ischemia and reperfusion injury. The American Journal of Surgery 154 , 211 to 218.
Taiwo, Y.O. & Levine, J.D. (1989). Prostaglandin effects after elimination of indirect hyperalgesic mechanisms in the skin of the rat. Brain Research 492 , 397 to 399.
Tillman, L.J. & Cummings, G.S. (1992). Biologic mechanisms of connective tissue mutability. In : Currier, D.P. & Nelson, R.M. (Eds.) (1992). Dynamics of Human Biologic Tissues (pp1-44). Philadelphia : F.A. Davis Company.
Takahashi, K., Olmarker, K., Holm, S., Porter, R.W. & Rydevik, B. (1993). Double level cauda equina compression. An experimental study with continuous monitoring of intraneural blood flow in the porcine cauda equina. Journal of Orthopedic Research 11 (1), 104 to 109.
Turvey, M.T., Fitch, H.L., Tuller, B., (1978a). Part V : degrees of freedom, co-ordinative structures and tuning. In : Kelso, J.A.S. (Ed). Human Motor Behavior : An introduction . Erlbaum. New Jersey.
Turvey, M.T., Shaw, R.E., Mace, W.M., (1978b). Issues in the theory of action : Degrees of freedom, co-ordinative structures and coalitions. In : Requin, J., (Ed). Attention and Performance VII . (557-595). Erlbaum. New Jersey.
Turvey, M.T., Shaw, R.E., Reed, E.S., Mace, W.M., (1981). Ecological laws for perceiving and acting : A reply to Fodor and Pylyshyn. Cognition , 9 , 237 to 304.
Vicenzino, B., Collins, D., Wright, A., (1996). The initial effects of a cervical spine manipulative physiotherapy treatment on the pain and dysfunction of lateral epicondylalgia. Pain , 68 , 1, 69 to 74.
Vicenzino, B., Wright, A., (1995). The effects of a novel manipulative physiotherapy technique on tennis elbow : a single case study. Manual Therapy , 1 , 30 to 35
Wall, P.D. (1995). Treatment of pain. In : Shacklock, M., (Ed). Moving in on Pain . Adelaide.
Warner, J.J.P., Micheli, L.J., Arslanian, L.E., Kennedy, J., Kennedy, R., (1990). Patterns of flexibility and strength in normal shoulders and shoulders with instability and impingement. The American Journal of Sports Medicine , 18 , 4, 366 to 375.
Whiting, H.T.A., (1980). Dimensions of control in motor learning. In : Stelmach, G.E., Requin, J., (Eds). Tutorials in Motor Behavior .(537-550). Nth Holland Pub. Co..
Williams, T.J. & Hellewell, P.G. (1992). Endothelial cell biology. Adhesions molecules involved in the microvascular inflammatory response. American Review of Respiratory Disease 146 , S45 to 50.
Woolf, C.J. (1983). Evidence for a central component of post-injury pain hypersensitivity. Nature 306 , 686 to 688.
Woolf, C.J. (1984). Long-term alterations in the excitability of the flexion reflex produced by peripheral tissue injury in the chronic decerebrate rat. Pain 18 , 325 to 343.
Woolf, C.J. & McMahon, S.B. (1985). Injury-induced plasticity of the fexor reflex in chronic decerebrate rats. Neuroscience 16 , 395 to 404.
Woolf, C.J., Shortland, P. & Sivilotti, L.G. (1994). Sensitization of high mechanothreshold superficial dorsal horn and flexor motor neurones following chemosensitive primary afferent activation. Pain 58 , 141 to 155.
Woolf, C.J. & Swett, J.E. (1984). The cutaneous contribution to the hamstring flexor reflex in the rat : an electrophysiological and anatomical study. Brain Research 303 , 299 to 312.
Woolf, C.J. & Wall, P.D. (1986). Relative effectivenss of C primary afferent fibres in evoking a prolonged facilitation of the flexor reflex in the rat. The Journal of Neuroscience 6 , 1433 to 1442.
Wright, A, Vicenzino, B., (1995). Cervical mobilisation techniques, sympathetic nervous system effects and their relationship to analgesia. In : Shaklock, M., (Ed), (1995). Moving in on Pain , 164 to 173. Butterworth Heinemann, Adelaide, Australia.
Zochodne, D.W. (1993). Epineural peptides : A role in neuropathic pain? Canadian Journal of Neurological Sciences 20 (1), 69 to 72.
Last update : 20 April 2008
 What We do
What We do What We Treat
What We Treat